A Vision for Continental-Scale Biology: Research Across Multiple Scales (2025)
Chapter: 2 Themes for a Continental-Scale Biology
2
Themes for a Continental-Scale Biology
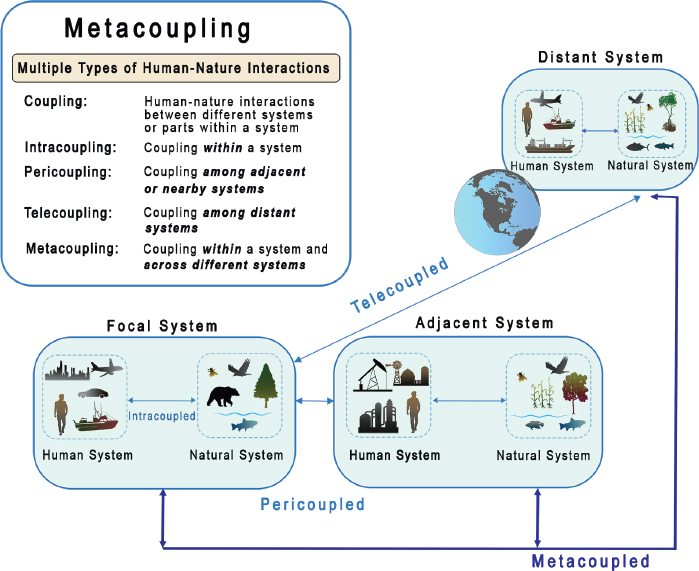
The committee identified four major themes in the types of research that CSB is particularly well positioned to support and in each theme, identified a series of research questions that could serve as pilots for implementing research projects. These themes and questions are neither mutually exclusive nor comprehensive, but rather are intended to stimulate discussion about directions for the development of the field (Figure 2-1). Although these themes have been extensively studied, previous research tended to focus on one particular scale. Thus, comparisons and interactions among different scales have not received sufficient attention. Furthermore, the literature on these themes is largely from biological and ecological perspectives. CSB aims to systematically and explicitly expand from a single scale to multiple scales, and from biological and ecological perspectives to integrated biological, ecological, and socioeconomic perspectives to account for pervasive human–nature interactions.
The four major research themes are interrelated, as shown in Figure 2-1. For instance, the fourth theme, “‘sustainability of ecosystem services’” is related to the first three because biodiversity and ecosystem functions are essential to produce ecosystem services, resilience and vulnerability are key in maintaining ecosystem services, and connectivity is central to shaping distribution and use of ecosystem services across space and over time.
Understanding scalability and how biological properties (e.g., patterns and processes) vary or remain the same across scales is an important area of inquiry for CSB. While different themes may have specific questions, a common question for all themes is how different biological properties vary or do not vary across spatial, organizational, and temporal scales. Furthermore, addressing all themes can benefit from a systems approach.

SOURCE: Stacy Jannis.
BIODIVERSITY AND ECOSYSTEM FUNCTION
A pivotal theme of ecological research over the past half century has been the relationship between biological variation, or biodiversity (in terms of taxonomy, function, phenotype, genotype or phylogenetic placement) and ecosystem functions, for example, the cycling of carbon, nitrogen, phosphorus, sulfur, oxygen, water, and trophic interactions.
Much of the experimental work has been conducted at local scales, but generalities have emerged to indicate that biodiversity is important to ecosystem function at multiple scales. As yet, there is scant evidence available to define these relationships and to decipher how they may change across scales—information that will be central to determining how biodiversity across spatial and temporal scales drives local to Earth system functions. An example comes from recent work to catalog Earth’s microbial biodiversity to better understand the function and structure of microbial communities that influence processes across entire ecosystems, from the gut microbes that help digest food to the microbes that help cycle carbon and nitrogen (see Box 2-1).
Ecosystem studies have tended to focus on collocated measures of biodiversity and ecosystem processes, such as the Jena experiment in Germany1 and the Cedar Creek experiment in the United States.2 CSB goes beyond typical ecosystem studies
___________________
1 See https://the-jena-experiment.de (accessed August 16, 2024).
2 See https://cedarcreek.umn.edu/about-cedar-creek-lter (accessed August 16, 2024).
BOX 2-1
Cataloging Earth’s Microbial Biodiversity Across Multiple Habitats
The majority of Earth’s biodiversity is microbial, and increasingly, scientists are realizing that microbes (bacteria, archaea, protists, fungi, and viruses) are the drivers of key processes in plants, animals, and across entire ecosystems, from the gut microbes that help digest food to the microbes that help cycle carbon and nitrogen. Understanding these processes will require new knowledge of the organization, evolution, functions, and interactions among the billions of the planet’s microbial species. Yet most microbes remain unknown to humans: a 2016 paper in Proceedings of the National Academy of Sciences (Locey and Lennon 2016) estimated that Earth could contain 1 trillion microbial genotypes, with only one-thousandth of 1 percent identified. The Earth Microbiome Project,a founded in 2010, seeks to catalog Earth’s microbial biodiversity by identifying and classifying microbes and identifying the factors that shape microbial communities.
The project leverages the power of crowd sourcing, relying on thousands of researchers around the world to collect samples from organisms and environments as diverse as the human gut, a bird’s mouth, the soil of an Antarctic valley, a lake in Germany, and the bottom of the Pacific Ocean. In a 2017 Nature publication, Thompson et al. (2017) detailed protocols, analytical methods, and software developed to process the 200,000 samples collected. The team decided to focus on bacteria, sequencing a 90-base-pair region of the 16S ribosomal RNA gene to act as a unique identifier of each species present in the sample.
The data generated have yielded new insights about the organizing principles that govern the structure of microbial communities. For example, Thompson et al. (2017) compared samples taken from different environments, finding that salinity and host association (e.g., the microbes were found in plants and animals) play significant roles as environmental factors that differentiate microbial communities.
In a 2022 Nature Microbiology publication, the Earth Microbiome Project team expanded the analysis to identify the metabolites present in each sample (Shaffer et al. 2022). Microbes produce metabolites as a byproduct of the reactions they are carrying out. By identifying and quantifying the different metabolites in each sample, researchers can learn about microbes’ interactions and activities. Working with an additional 880 samples taken from sites around the world, the team used techniques including gas chromatography-mass spectrometry to study the metabolite profile of each sample. In addition to sequencing a region of the 16S ribosomal RNA gene, in this study, the team also sequenced regions of the 18S ribosomal RNA and internal transcribed spacer genes to serve as an identifier for protist and fungal species.
The team developed standardized protocols for each technique, which will make it easier for researchers to compare data from different studies to provide a global-scale view of microbial communities across multiple habitats. All the data generated by the Earth Microbiome Project are available on an open-source platform, and teams of researchers around the world continue to gain new insights from the vast dataset.
__________________
a See https://earthmicrobiome.org/ (accessed March 8, 2024).
by integrating collocated measures in the focal place, such as an experiment site, and measures of flows (of organisms, materials, information, water, etc.) between the focal place and other places (e.g., integration of movement ecology with ecosystem ecology). This integration requires new study designs that account for measures of flows and reciprocal effects of flows and internal factors as indicated in collocated measures.
A CSB approach offers the opportunity to use emerging tools, including satellite remote sensing, genetic sequencing, multi-omics, artificial intelligence, and automated monitoring systems, together with developing theory, to examine the linkages between biodiversity and ecosystem functions across scales, and to learn how these relationships may shift in the face of global change. The committee identified the following example research questions in this theme:
- How does complexity of a biological system at one scale influence emergent properties of the system at the next scale, and how do those properties feed back to influence complexity and variation at all scales?
- How do ecosystem functions emerge from biological complexity, what mechanisms are involved, and how do these vary across temporal and spatial scales? How do major global changes, including climate change, human-induced land- and water-use changes and the spread of pests and pathogens, impact how the diversity of life influences and interacts with functions and processes of emergent systems?
- How do human-induced changes in land use and water use, habitat fragmentation and, conversely, habitat connectivity at local and regional scales affect continental-scale biodiversity?
- How does theory support our understanding of the causes and consequences of continental-scale biodiversity in the context of major global changes and human activities?
- How does the distribution of composition, diversity, and complexity of ecosystems influence the cycling of carbon, nutrients, and water at continental scales, and how do these cycles feed back to influence the composition, diversity, and complexity of local systems?
- How do internal factors in a focal place and flows between focal and other places influence biodiversity and ecosystem functions across scales?
RESILIENCE AND VULNERABILITY
Resilience is the capacity of a system to withstand or recover from human and environmental disturbances3 (Pickett et al. 2013, Reid et al. 2014). Examples include climate change, pollution, habitat loss, and landscape fragmentation, which are impacting ecosystems all over the planet (Mann et al. 2008, Melillo et al. 2014). The effects of human and environmental disturbances can be identified as “press,” which means they are chronic, or “pulse,” which means they are acute (Inamine et al. 2022). Resilience to
___________________
3 See https://www.resalliance.org/resilience (accessed September 3, 2024); see also Levin (2024).
disturbances consists of three components: stability, resistance, and recovery. A stable ecosystem can resist disturbance by maintaining structure, function, and composition. A resilient ecosystem can recover from disturbances over time and essentially return to its original state. Ideally, multiple metrics should be used to provide a composite assessment (Dakos and Kéfi 2022). In contrast, vulnerability is the extent to which a system (or a system component) is likely to experience harm due to exposure to disturbances (Turner et al. 2003). A vulnerable system is sensitive to disturbances and lacks adaptive capacity when conditions change.
Much research on resilience and vulnerability in biological systems has been conducted at a particular scale. However, little evidence exists regarding how resilience, vulnerability, and their relationships may change across scales. Such information is critical to understanding sustainability and many other aspects of biological systems across different scales. A CSB approach offers the opportunity to study resilience, vulnerability, and their relationships across scales, and to examine how these relationships may vary under global change, human activities, and connectivity. CSB research can also be used to assess the sensitivity of biological systems and their adaptive capacity to resist, recover, or change in biological composition and function across multiple scales in response to disturbances.
Resilience and vulnerability are affected by factors not only within a place but also in adjacent and distant places. For example, population dynamics in a place are affected by not only internal factors such as birth rates but also interactions with adjacent and distant places such as migration. If the population in the focal place is eliminated or reduced by a disturbance, the population in the focal place may or may not recover without immigrants from adjacent and distant places. Such immigrants may be affected by human activities, such as human-assisted migration of species in response to the disturbance.
Understanding the factors that influence the resilience and vulnerability of individuals, populations, communities, and ecosystems across scales to changing conditions will be essential for informing multiscale strategies for the conservation of biodiversity and management of ecosystem services. One example comes from the research that explores how the microbial communities that make up coral reef biofilms could provide an indicator of the health and resilience of the reef ecosystem (see Box 2-2). Another describes the effects of environmental and human factors on vulnerability of native plant populations to invasive species (Box 2-3).
Critical questions remain to assess a system’s resilience and vulnerability that through drivers, processes or impacts are fundamentally linked to CSB:
- How do biodiversity and human activities influence resilience of biological systems at multiple spatial and temporal scales? How does this relationship shift across scales of biological organization, from the population (diversity within a species) to the biome?
- Which ecosystems and functions at what scales are most exposed and vulnerable to natural and human disturbances?
- What are the tipping points and thresholds of resilience and vulnerability across scales? What continental-scale drivers affect the spatial and temporal patterns of
- ecosystem vulnerability at local, regional, and national scales, and how do local to regional drivers affect vulnerability and resilience at the continental scale?
- How does the resilience or vulnerability of one system—for example, to land use or climate change—affect resilience or vulnerability of other systems near and far?
CONNECTIVITY
Ecosystems do not exist in isolation. The world is more connected due to a variety of factors such as globalization, climate change, land and water use, disease spread, and environmental changes. Such connections have important biological and socioeconomic consequences across multiple scales, including the continental scale. Increasingly, scientists are understanding that human–nature interactions or natural events in one place can have profound effects on ecosystems in other places, both nearby and distant. For example, deforestation may be facilitated by adjacent roads (Cropper et al. 2001) and distant demand for agricultural products (DeFries et al. 2010). Wildfires have increased in many places (MacDonald et al. 2023) and can influence air quality in areas far from the actual fires (Jaffe et al. 2020); forest wildfires in Oregon have affected air quality on the East Coast of the United States (Miller, 2021), and the Canadian wildfires in 2023 caused air quality alerts as far south as the Carolinas (Dennis and Koh, 2023).
Many studies have examined linkages in time and space. Examples include the stream continuum concept (Stallard 1998), transoceanic transport of nutrients (Vitousek 2004), and seabird movement of pelagic nutrients into land and intertidal rocky shores (Healing et al. 2024). However, more systematic approaches are needed to effectively integrate biota, habitats, ecosystem functions and materials, and humans across time and space. For example, while spatial connections in metapopulations (and metacommunities and metaecosystems) have been extensively studied (Holt 1997, Leibold et al. 2004, Loreau et al. 2003), their main focus has been on ecological dimensions with little explicit attention to human dimensions. Furthermore, previous studies usually do not provide an explicit way of contrasting spatial linkages (e.g., adjacent and distant). The explicit integration can help scientists understand, avoid, and solve problems when, for example, sustainability efforts focus on one ecosystem issue without realizing how that may impact connected systems near and far.
CSB can offer tools that can combine data and research from various realms across time and space, helping to build a stronger understanding of the system as a whole. For example, interdisciplinary frameworks provide systematic ways to combine data and insights from different ecosystems across time and space. A number of frameworks have been developed to address multiscale and cross-scale issues (e.g., Allen and Starr 1982, Folke et al. 2011, Heffernan et al. 2014, Levin 1992, Peters et al. 2008, Rose et al. 2016). They have provided useful insights and inspired research efforts on CSB. However, they do not explicitly specify human–nature interactions and feedbacks within as well as between adjacent and distant systems, which are widely recognized common real-world phenomena at multiple scales.
BOX 2-2
Biodiversity in Coral’s Slimy Biofilms as an Indicator of Reef Resilience
Coral reefs harbor millions of bacteria, both floating free in the water and forming a slimy reef coating called biofilm. Understanding the mix of microbes that make up coral biofilm could provide crucial insights on the ecosystem services they provide—and according to recent research, could also provide a measure of the resilience of the reef ecosystem (Remple et al. 2021).
Biofilm is a bacterial matrix that coats surfaces—in this case, coral reefs. The microbes that make up biofilm keep coral reefs healthy: biofilm bacteria help with organic matter decomposition, nutrient cycling, and recruiting the next generations of coral larvae to sustain reef structure. These ecosystem services are likely governed by the composition of the biofilm, a delicate balance of different species of bacteria that depends on environmental conditions, including factors such as fluid dynamics and nutrient availability.
Until recently, few studies have focused on the factors that influence coral biofilm composition. More is known about free-floating marine microbes. For instance, research has shown that ocean acidification and warming can cause a decrease in microbial diversity in some coral ecosystems, and other studies have shown that nutrient pollution can boost populations of disease-causing bacteria.
In combination with other stressors such as overfishing, nutrient pollution also can boost the growth of fleshy seaweeds called macroalgae in coastal ecosystems. Once the macroalgae starts to grow, it can launch a feedback loop that shifts the ecosystem from coral dominated reefs to those dominated by algae. The exact mechanisms of this are still being worked out, but differences in the metabolic processes carried out by macroalgae and corals lead to differences in the composition of the organic matter they exude into the water column. Macroalgae in the ecosystem change the physical and chemical qualities of the water, which restructures microbial communities and favors a less diverse mix of microbial species that is less able to support coral vitality.
Remple et al. (2021) investigated how nutrient pollution and algae impact the composition of coral biofilm. The team cultured biofilms in environments containing sand, algae, or corals and with different levels of added nutrients, and surveyed the bacterial communities in each biofilm sample.
The study found that at earlier stages, biofilms were dominated by “metabolic generalists”—microbes that are less specialized in the use of organic carbon compounds. Over time, biofilm communities became more diverse and contained markedly different microbial communities than the free-floating bacteria within the same samples. Adding nutrients decreased the diversity of biofilm communities, favoring primary producers and bacteria that thrive in nutrient-rich conditions rather than the nutrient-cycling bacteria typically found in biofilms.

SOURCE: Stacy Jannis.
Biofilms cultured with algae were less diverse than those grown with corals, and were dominated by microbial species adept at growing quickly on nutrient-rich carbon compounds. In contrast, the biofilms cultured with corals maintained a rich diversity of microbial life throughout the experiment, although the addition of nutrients caused a decrease in diversity in mature biofilms. This indicates that the addition of nutrients could derail microbial diversity even in coral-dominated reefs.
Finding that distinct differences exist between the composition of biofilms in coral-dominated reefs and those that include algal species paves the way for future understanding of how shifts in coral ecosystems may impact key microbial processes that are crucial for reef resilience.
Biofilm biodiversity could also potentially provide an indicator of ecosystem health: a more resilient ecosystem may be able to maintain biofilm diversity even as environmental conditions change, and conversely, a loss of biofilm diversity could indicate that environmental conditions are decreasing ecosystem function.
BOX 2-3
Effects of Local, Landscape, and Regional Drivers of Vulnerability to Plant Invasions
Leveraging multiscale data on native and non-native plant species across National Ecological Observatory Network sites, Ibáñez et al. (2023) quantified the compound effects of local, landscape, and regional drivers of vulnerability to plant invasions. Vulnerability was influenced by temperature, precipitation, productivity, and human modification, but the magnitude and nature of these influences varied widely across ecoregions. For example, in colder regions, native species were more vulnerable to local-scale losses, probably due to shorter growing seasons favoring invasive plants, whereas in warmer regions, the impact of non-natives was more pronounced at landscape scales, suggesting that local non-native effects can extend over larger areas. By accounting for cross-scale effects, the study provided a nuanced view of vulnerability that could aid in localized and broader-scale decision making for managing plant invasions. The results also underscore the importance of considering multiple scales and environmental factors to better predict and manage the impacts of invasive species on native plant communities.
A recent framework that built upon, expanded from, and integrates various previous frameworks is the metacoupling framework (Liu 2023), which explicitly considers the interactions that take place within a system (including human–nature and nature–nature interactions) and interactions with adjacent and distant systems. Nature includes both biotic components (animals, plants, and microorganisms) and abiotic components (e.g., air, climate, geophysical conditions, soil/land, water). Humans consist of social, economic, political, cultural, and other components. The framework takes a systems approach (e.g., by treating each place as a coupled human and natural system, and all interacting places near and far as metacoupled human and natural systems). It integrates social, economic, and ecological connections and feedback within and between systems and provides a way to enhance synergies and reduce trade-offs between them. This framework has been applied at multiple scales ranging from local and regional scales (Wang et al. 2024, Zhao et al. 2021) to national and global scales (Carlson et al. 2020, Xiao et al. 2024, Xu et al. 2020). It is discussed in more detail in Box 2-4, which illustrates how the metacoupling framework can help scientists and policymakers integrate data on the connections between humans and natural ecosystems, and as an example, to inform fisheries management decisions.
As mentioned above, biodiversity, ecosystem functions, resilience, and vulnerability are affected by not only internal factors within a focal place but also factors in adjacent and distant places. The metacoupling framework accounts for all these factors.
It goes beyond the biological or ecological approaches because the movement of species and materials, for example, across boundaries is not just driven by natural factors, but also driven by human factors and human–nature interactions. For example, the materials that move across the landscapes may be released by human activities such as agricultural practices including fertilizer applications. Increasingly, migration of many species is affected by humans.
Currently, the metacoupling approach is limited by a lack of understanding of the rate and quantity of connections and feedback between systems that influence these dynamics. To truly deliver on the promise of metacoupling analysis requires both novel and robust methods of data integration and development and refinement of theory to support such advancements. Research questions to support these goals include:
- How do human–nature interactions vary across organizational, spatial, and temporal scales, and what are the impacts of economic globalization and environmental changes on these dynamics?
- How do human activities and environmental changes (including climate change) in one system generate cascading effects across adjacent and distant systems?
- How do interactions within a focal system, between adjacent systems, and between distant systems create feedback, synergistic, or trade-off effects on resilience and sustainability of biodiversity and ecosystems across scales?
- How do risks associated with invasive species, pathogen emergence, and other shocks such as natural disasters spread across adjacent and distant systems at multiple scales?
SUSTAINABILITY OF ECOSYSTEM SERVICES
Ecosystem services are the benefits people obtain from ecosystems, including the provisioning of food and water; the regulation of floods, drought, land degradation, and disease; nutrient cycling and soil formation; and nonmaterial benefits such as recreational, spiritual, and religious benefits of nature (Costanza et al. 1997, Daily and Matson 2008, Diaz et al. 2019).
Sustainability relates to an ecosystem’s ability to continue to provide services, without a substantial reduction in the level of ecosystem services, in the face of environmental and human disturbances or state changes such as land changes due to urbanization (Grimm et al. 2008, Seto et al. 2012, Shepherd et al. 2002, Turner et al. 2007). Being able to assess the sustainability of an ecosystem service across scales is critical for developing targeted multiscale mitigation and adaptation strategies for sustainability. However, previous research and management tended to largely focus on sustainability of ecosystem services at a single scale (e.g., Ashrafi et al. 2022) although there is a strong need for pursuing multiscale management and sustainability (e.g., Clark and Harley 2020, Hou et al. 2023, Tavárez et al. 2022).
CSB can contribute to our understanding of sustainability by examining the processes through which natural systems give rise to ecosystem services across scales, and in turn contribute to inclusive wealth. Inclusive wealth encompasses all the assets
BOX 2-4
A Metacoupling Framework to Help Optimize Salmonid Research, Management, and Policymaking
Salmonids—fish in the salmon family including salmon, trout, char, and freshwater whitefish—are not only key species in their ecosystems, playing roles as both predators and prey, in nutrient cycling, and as indicators of ecosystem impairment, but are also economically and culturally important as a source of food for people and a target for recreational fishing. Increasingly, anthropogenic factors such as climate change, groundwater withdrawal, and habitat disruption are impacting salmonid populations.
Understanding how these factors interact is important for monitoring the health of salmonid fisheries, but to date most fisheries research does not have a systematic way to integrate the influences of biota, habitats, and humans across time and space. Instead, most sustainability efforts have solved problems one at a time, often at the cost of other components of the system.
Recent advances in theory and methods have helped develop a “metacoupling” approach to studies of coupled human and natural systems (CHANS) that could offer insights for salmonid management and policymaking. The metacoupling framework would provide a useful tool for studying the interactions between humans and nature that occur within a place (in this example, the salmonid fisheries), and between adjacent places and distant places, allowing the analyses to scale from local to global levels.
Overall, the metacoupling framework seeks to integrate social and ecological information, characterize trade-offs and feedbacks across scales, and understand the roles that different members of the community play, including anglers, landowners, and people who work in the recreational fishing industry. The ultimate goal is a holistic, “big picture” view that informs fishery management decisions to improve the relationships between human and natural systems as a whole, rather than in specific physical places in isolation, to better sustain salmonid fisheries locally, regionally, and globally.
This metacoupling approach was illustrated by Carlson et al. (2020), which analyzed brook char and brown trout fisheries in two adjacent cold-water streams in Michigan, the Twin and Chippewa creeks. These salmonid species have enormous socioeconomic and recreational significance in the region, representing a crucial component of statewide fisheries that generate $4.2 billion in overall economic effect. At the same time, there are concerns that groundwater withdrawal to supply a drinking water bottling plant could alter stream hydrology, which has caused controversy among landowners and anglers over how the hydrologic changes may impact salmonid populations.
Using the metacoupling framework, the study authors combined information on how water, information, fish, people, and money are moving through individual CHANS (such as Twin Creek), between adjacent CHANS (such as Twin and Chippewa creeks), and, at the regional level, through non-adjacent CHANS (such as Twin Creek and Hoffman Creek).
One insight from this analysis is that although statewide regulation on water withdrawals is effective, local community members strongly prefer local-scale governance because it engages local communities and considers the groundwater dynamics of individual streams and watersheds.
Another insight was about a proposed transition from riparian forest to shrubs on the banks of the Twin and Chippewa Creeks. This transition could decrease water temperatures by 0.09°C, helping to maintain the cold-water habitat needed for the brown trout and brook char. However, the metacoupling analysis indicated that this land-use change would reduce angling, and the associated revenue streams for local communities, because the natural beauty of the forested streams is highly valued by salmonid anglers and is a key factor in decisions about which stream to visit, underscoring that what may be ecologically beneficial for the fishery may be socially and economically detrimental, and vice versa. This perspective could help fishery managers to identify a holistic solution that balances social, economic, and ecological trade-offs for sustainable salmonid management.
SOURCE: Stacy Jannis.
that contribute to human well-being—including the natural capital, human capital, and produced capital—and is a measure designed to address whether society is on a sustainable trajectory (Polasky et al. 2015, UNEP 2018). Sustainability can then be defined as nondeclining human well-being, based on metrics of inclusive wealth.
CSB research brings knowledge, via systems’ resilience and vulnerability to stressors, on how sustainability of ecosystem services may be affected across organizational, temporal, and spatial scales. For example, trees offer crucial ecosystem services, such as carbon sequestration, air pollution removal, and wood production. Such services vary among different tree species and lineages and different regions of the contiguous United States (Cavender-Bares et al. 2022). Box 2-5 discusses how the ecosystem service of slope stability changes across spatial scales with changing rainfall and species composition, and Box 2-6 describes how tree fecundity affects the long-term maintenance of ecosystem services provided by forested areas.
The committee identified a series of research questions to explore how CSB can increase understanding of sustainability, including:
- Which ecosystem services are most sustainable at different scales in the face of human activities and environmental change?
- How do socioeconomic, urbanization, and land-use change affect ecosystem services across local to continental scales?
- What factors across scales affect ecosystem services at the scale of interest, and how does the sustainability of an ecosystem service shift across spatial scales?
- How does sustainability of ecosystem services at one scale affect sustainability at other scales?
EXAMPLES OF INTERWORKING OF THE FOUR THEMES
Everglades Restoration
Florida’s Everglades have been described as a river of grass, formed as waters from Lake Okeechobee flowed slowly southward over sawgrass marshes, cypress swamps, wet prairies, and other habitats before reaching Everglades National Park and Florida Bay. The water that created the unique ecosystem, however, also posed a flooding risk for people. In 1928, the deadly Okeechobee Hurricane sent 15-foot waves from the lake, killing more than 2,500 people. In 1947, the U.S. Army Corps of Engineers (the Corps) proposed the Central and South Florida Project to provide flood protection for growing urban development and agriculture activities in South Florida. By 1960, the Corps had built 720 miles of canals and 1,000 miles of levees that profoundly altered the region’s wetlands. Today, at 1.5 million acres,4 the Everglades is half its original size and is impaired by contaminated runoff from cities and farms (NASEM 2023).
Since the early 1990s, a coalition of local, state, and federal agencies, nongovernmental organizations, local tribes, and citizens has been working to reverse the
___________________
4 See https://www.nps.gov/ever/planyourvisit/basicinfo.htm (accessed August 30, 2024).
BOX 2-5
Bridging Spatial Scales: How the Ecosystem Service of Slope Stability Changes with Rainfall Amounts and Forest Species Composition
Bridging spatial scales, from belowground roots to hillslopes, has allowed research to inform how the ecosystem service of slope stability changes with rainfall amounts and forest species composition. Roots reinforce soils against shallow landslide initiation by increasing apparent cohesion. The magnitude of the reinforcement provided by roots depends on the number, the size distribution, and the elastic properties of roots that cross potential failure planes. In forested landscapes, the variability in belowground properties at a hillslope scale makes it challenging to predict root reinforcement at the landscape scale. Topography can affect root strength through changes in root cellulose content, with stronger roots on divergent landscape positions and weaker roots on convergent landscape positions (Hales et al. 2009). Using an ecohydrologic model fused with a landslide model allowed spatial scaling to generate factor-of-safety maps that predict landslide potentials (Band et al. 2012). Further scaling relationships between tree height, root biomass, and dynamic soil moisture increased the model’s predictive ability (Hwang et al. 2015). Ultimate model refinements incorporated a demonstrated feed-forward response of root strength to soil moisture; as roots gain moisture content, they become weaker (Hales and Miniat 2017). Landslide maps for counties in western North Carolina were used to develop steep-slope ordinances such that development could not occur on steep slopes with a high likelihood of failure. This work has also been influential in coordinating with the National Weather Service to establish guidelines for when they include wording on landslide hazards in their public advisories for floods and flash floods in the region.
damage to the Everglades. Led by the U.S. Army Corps of Engineers (USACE), and the South Florida Water Management District, the blueprint for the restoration effort, the Comprehensive Everglades Restoration Plan (CERP), was published in 1999. The plan proposed 68 individual projects to be constructed over an estimated 30 to 40 years in the South Florida Ecosystem. CERP’s primary goal is to “get the water right”—that is, to deliver the right amount and quality of water to restore characteristics of the historic ecosystem (NASEM 2023). The plans included the creation of below- and aboveground water storage, water quality treatment, and the removal of barriers to the historic sheet flows of water (Figure 2-2).
CERP is a good illustration of the interplay of the four themes laid out in this report, from local to ecosystem scales. At the center is the desire to restore and protect long-term sustainability of the South Florida ecosystem and the services it provides. Wetland ecosystems filter pollutants, excess nutrients, and sediments from the water,
BOX 2-6
Effect of Tree Fecundity on the Long-Term Maintenance of Ecosystem Services Provided by Forested Areas
The long-term maintenance of ecosystem services provided by forested areas will be highly dependent on the functionality of these systems, that is, the maintenance of healthy ecosystems (Turner and Daily 2008). Tree reproduction rates that keep pace with current environmental changes will be critical for the preservation of these ecosystems, either by maintaining populations in place or by tracking climate change via migration (Davis and Shaw 2001, Martinez-Vilalta and Lloret, 2016). Linking locally collected records of tree fecundity across North America, Clark et al. (2021) built a continental Masting Inference and Forecasting (MASTIF) network of primary data to quantify tree species fecundity across North America. They describe a biogeographic divide between the East, increasing fecundity, and the West, decreasing fecundity. These continental-scale forest responses are driven by both the direct and indirect effects of climate change on individuals via stand-level features. This analysis also shows how, despite drier summers, hot spots of tree fecundity resulted from the interaction between regional climatic trends, spring rising temperatures and declining moisture deficits, and optimal local stand structure that affected individual tree growth. The potential effects of changing environmental conditions on tree fecundity were the result of drivers acting and interacting across spatial scales.
which helps maintain water quality. The wetlands also replenish aquifers that serve the drinking water needs for one-third of Floridians and irrigation for much of the state’s agriculture. In addition, the national parks and wildlife refuges offer opportunities for tourism and enhancing health and well-being that people obtain from enjoying nature.
Achieving sustainability in the Everglades requires a high degree of integration of scientific knowledge about the biodiversity of the system. The Everglades are home to more than 360 bird species, 60 reptile species, and 40 mammal species, and a wide variety of plant life, including bromeliads, cacti, succulents, native grasses, seagrasses, and wildflowers. Longstanding CERP challenges include balancing the right quantities of water and timing of flows to habitats hosting the endangered Cape Sable seaside sparrow, providing food and nesting space for the endangered snail kite and wading bird species, and addressing degradation of tree islands and ridge and slough topography by water levels that were too high in some places and too low in others.
A big part of restoration decision making involves consideration of connectivity in the system, including trade-offs, interactions, and synergies across multiple scales in the system from organisms to habitats, to the whole ecosystem and adjacent ecosystems. CERP hydrological and ecological modeling and monitoring efforts strive to examine
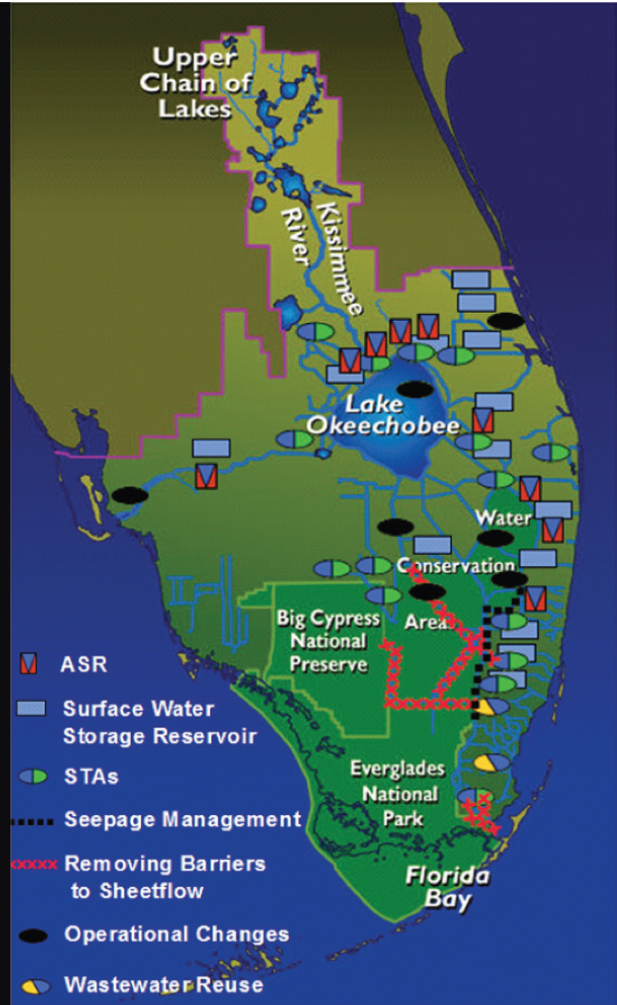
the effects of restoration on these diverse system components. Many of the CERP projects address the connectivity that has been lost between parts of the system, and how actions taken in one part of the ecosystem influence conditions in other parts of the system. For example, a large project aims to increase the duration and magnitude of flows through the central Everglades into the northeast portion of Everglades National Park. However, increasing flows, even at levels that meet current water quality criteria, have been shown to adversely impact the biota, with losses of periphyton and increasing cattails evident in pilot testing. Thus, planners are grappling with ways to mitigate water quality impacts while maximizing flow benefits.
Finally, CERP projects are increasingly focused on resilience to expected future change, mostly as a result of climate change and sea-level rise. Impacts of sea-level rise can include flooding, shifts in extent and distribution of wetlands and other coastal habitats, and salinity intrusion into estuaries and groundwater systems. Increasing air temperatures reduce runoff and impact water availability, unless precipitation increases to counter these effects. CERP projects such as the South Florida Water Management District’s new Sea Level Rise and Flood Resiliency Plan proposes infrastructure and nature-based solutions to address vulnerabilities to sea-level rise, storm surge, and extreme rainfall events.
Pandas, People, and Policies
This example encompasses all four themes presented above, taking a systems approach and using Wolong Nature Reserve in southwestern China as an illustration of terrestrial systems (Figure 2-3). The reserve was established in 1975 to conserve giant pandas, a global wildlife icon (Liu et al. 2016). It is within a global biodiversity hotspot (Myers et al. 2000). With an area of 2,000 km2, it is home to several thousand plant and animal species, including approximately 150 (or 8% of the total population) wild giant pandas, and several thousand local residents (mainly farmers). Wolong constitutes a typical coupled human and natural system, where humans affect the natural systems (including panda individuals and habitat), which provide a variety of ecosystem services, such as water purification, carbon sequestration, and food (Yang et al. 2013). Wolong is vulnerable but is also resilient to disturbances such as the 2008 Wenchuan earthquake (Zhang et al. 2014). It has been a living laboratory of systems integration for understanding and managing coupled human and natural systems, including biological systems underpinning CSB, since 1996 (Liu et al. 2016).
Scaling has been a major research topic in Wolong and beyond. The spatial patterns of panda habitat based on data from satellites and field surveys vary at different scales. At a local scale (e.g., forest-stand patch), an entire area may be suitable or unsuitable. At a large scale, some areas are suitable habitats while other areas are unsuitable. The discovery in Wolong of faster growth in the number of households (a major driver behind biodiversity and habitat dynamics) than human population size led to the finding of a similar pattern at the global scale (Liu et al. 2003).
There are different or similar patterns across temporal scales. Using GPS (global positioning systems) collars on individual wild giant pandas, Zhang et al. (2015) inves-

SOURCE: Andrés Viña and Jianguo Liu.
tigated the daily and seasonal activity patterns. Contrary to the literature, most pandas were not crepuscular but showed three daily activity peaks, in the morning, afternoon, and around midnight. In terms of seasonal pattern, panda activity peaked in June, decreased in August and September, and increased again from November to March of the next year. For the total amount of panda habitat, the general temporal patterns of loss and recovery in 1976–2013 are similar in Wolong Nature Reserve (Liu et al. 2016) and across the entire geographic distribution range scale (Xu et al. 2017).
Emergent properties appear at higher organizational scales. Connor et al. (2023) used noninvasive fecal genetic (DNA) sampling to observe panda individuals in a wild population and infer association networks according to their spatiotemporal patterns. Even though the panda was thought of as a solitary species, the social network analyses revealed that cluster members preferred to associate with each other. This social clustering is an emergent property that does not show at the individual or DNA scales (Figure 2-3). These results suggest that many other “solitary” species may also have strong associations among individuals and emergent properties at higher organizational scales.
Multiscale interactions and feedback among pandas, people, and policies shape systems dynamics, according to the systems approach that integrates various sources of data (e.g., remote sensing, field investigations, tracking through global positioning system collars, socioeconomic surveys, government documents) and a portfolio of methods (e.g., DNA analysis, social network analysis, statistical analysis, systems modeling, agent-based modeling). Results indicate that humans affect panda habitat through activities such as farming and forest harvesting. Changes in panda habitat prompt the government to develop and implement new policies, and new policies change human activities, which in turn affect panda habitat across different scales (Liu et al. 2016).
Research on Wolong inspired the development of the metacoupling framework that integrates different types of human–nature interactions across space. From this perspective, Wolong is a focal system and interacts with adjacent systems such as neighboring counties where some women marry men in Wolong and move into Wolong, and with distant systems such as countries like the United States where many people visit Wolong as tourists. Conversely, Wolong may affect nearby areas where wild pandas move out of the reserve and distant areas where pandas in Wolong’s breeding center are loaned to zoos such as the San Diego Zoo and the Smithsonian National Zoo in the United States.
Conservation efforts across local to global scales have led to the transition to sustainability. At the local scale, farmers in Wolong return cropland to forests, and reserve staff implement and monitor conservation efforts. Provincial and central governments, as well as international organizations on continents such as Asia, Europe, and North America, provide financial and technical support for the local farmers and reserve staff. Some members of the international and interdisciplinary team have also written blogs about the research for the general public. Science communicators and global news media outlets (e.g., BBC, The New York Times, Time for Kids, China’s Xinhua News Agency) have widely reported the findings. These and other efforts have greatly enhanced stakeholder engagement as well as policies and practices. As a result, the collective action has transformed the habitat from long-term losses (even after the establishment of Wolong as a nature reserve) to recovery and the ultimate removal of the panda from
the endangered species list of the International Union for Conservation of Nature in 2016. In this sense, panda habitat and population, as well as many ecosystem services, such as carbon sequestration, are on the trajectory to sustainability.
Methods and insights from Wolong have been applied to understanding and managing biodiversity as well as ecosystem services and other environmental challenges beyond Wolong, such as the Qionglai Mountain Range, Panda Geographic Range in China, other parts of Asia, and many other parts of the world (Figure 2-3). For example, the agent-based model developed for Wolong has been adapted to Chitwan National Park in Nepal (An et al. 2014). The habitat mapping methods developed in Wolong enabled studies on giant panda habitat dynamics across the species’ geographic range (Viña et al. 2010). The work in Wolong also inspired efforts to detect changes in protected areas at the national, continental, and global scales (Yang et al. 2019, 2021). Comparisons between Wolong and other areas with different socioeconomic–ecological conditions around the world indicate that they share many complex attributes (e.g., time lags, nonlinearity, legacy effects, heterogeneity) (Liu et al. 2007).
The broad utilities of multiscale research on the four integrated themes demonstrated in this example suggest their potential value for studying other sites. These include sites of the Long-Term Ecological Research, International Long-Term Ecological Research, Long-Term Agroecosystem Research to NEON, as described in Chapter 4.
REFERENCES
Allen, T.F.H., and T.B. Starr. 1982. Hierarchy: Perspectives for Ecological Complexity. Chicago: University of Chicago Press.
An, L., A. Zvoleff, J. Liu, and W. Axinn. 2014. Agent-based modeling in coupled human and natural systems (CHANS): Lessons from a comparative analysis. Annals of the Association of American Geographers 104: 723-745. https://doi.org/10.1080/00045608.2014.910085.
Ashrafi, S., R. Kerachian, P. Pourmoghim, M. Behboudian, and K. Motlaghzadeh. 2022. Evaluating and improving the sustainability of ecosystem services in river basins under climate change. Science of the Total Environment 806 https://doi.org/10.1016/j.scitotenv.2021.150702.
Band, L.E., T. Hwang, T.C. Hales, J.M. Vose, and C.R. Ford. 2012. Ecosystem processes at the watershed scale: Mapping and modeling ecohydrological controls of landslides. Geomorphology 137:159-167. doi:10.1016/j.geomorph.2011.06.025.
Carlson, A.K., W.W. Taylor, and S.M. Hughes. 2020. The metacoupling framework informs stream salmonid management and governance. Frontiers in Environmental Science 8:27. doi:10.3389/fenvs.2020.00027.
Cavender-Bares, J.M., E. Nelson, J.E. Meireles, J.R. Lasky, D.A. Miteva, D.J. Nowak, W.D. Pearse, M.R. Helmus, A.E. Zanne, W.F. Fagan, C. Mihiar, N.Z. Muller, N.J.B. Kraft, and S. Polasky. 2022. The hidden value of trees: Quantifying the ecosystem services of tree lineages and their major threats across the contiguous US. PLOS Sustainability and Transformation 1(4):e0000010. https://doi.org/10.1371/journal.pstr.0000010.
Clark, W.C. and A. Harley. 2020. Sustainability science: Toward a synthesis. Annual Review of Environment and Resources 45:331-386. https://doi.org/10.1146/annurev-environ-012420-043621.
Clark, J.S., R. Andrus, M. Aubry-Kientz, Y. Bergeron, M. Bogdziewicz, D.C. Bragg, D. Brock-way, et al. 2021. Continent-wide tree fecundity driven by indirect climate effects. Nature Communications 12:1242. https://doi.org/10.1038/s41467-020-20836-3.
Connor, T., K. Frank, M. Qiao, K. Scribner, J. Hou, J. Zhang, A. Wilson, V. Hull, R. Li, and J. Liu. 2023. Social network analysis uncovers hidden social complexity in giant pandas. Ursus (34e9):1-13. https://doi.org/10.2192/URSUS-D-22-00011.1.
Costanza, R., R. d’Arge, R. de Groot, S. Farber, M. Grasso, B. Hannon, K. Limburg, S. Naeem, R.V. O’Neill, J. Paruelo, R.G. Raskin, P. Sutton, and M. van den Belt. 1997. The value of the world’s ecosystem services and natural capital. Nature 387:253-260. https://doi.org/10.1038/387253a0.
Cropper, M., J. Puri, and C. Griffiths 2001. Predicting the location of deforestation: The role of roads and protected areas in North Thailand. Land Economics 77(2):172-186. https://doi.org/10.2307/3147088.
Daily, G., and P.A. Matson. 2008. Ecosystem services: From theory to implementation. Proceedings of the National Academy of Sciences of the United States of America 105:9455-9456. https://doi.org/10.1073/pnas.0804960105.
Dakos, V., and S. Kéfi. 2022. Ecological resilience: What to measure and how. Environmental Research Letters 17(4): 043003. https://doi.org/10.1088/1748-9326/ac5767.
Davis, M.B., and R.G. Shaw. 2001. Range shifts and adaptive responses to Quaternary climate change. Science 292:673-679. https://doi.org/10.1126/science.292.5517.67.
DeFries, R.S., T. Rudel, M. Uriarte, and M. Hansen. 2010. Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nature Geoscience 3(3):178-181. https://doi.org/10.1038/ngeo756.
Dennis, B., and J. Koh. 2023. Smoke from Canadian wildfires engulfs East Coast, upending daily life. The Washington Post. https://www.washingtonpost.com/climate-environment/2023/06/07/air-quality-nyc-us-canada-wildfire-smoke/ (accessed March 12, 2024).
Diaz, S., J. Settele, E.S. Brondízio, H.T. Ngo, J. Agard, A. Arneth, P. Balvanera, et al. 2019. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 366(6471):eaax3100. https://doi.org/10.1126/science.aax3100.
Folke, C., Å. Jansson, J. Rockström, P. Olsson, S.R. Carpenter, F.S. Chapin III, A.-S. Crépin, et al. 2011. Reconnecting to the biosphere. AMBIO 40:719-738. https://doi.org/10.1007/s13280-011-0184-y.
Grimm, N.B., D. Foster, P. Groffman, J.M. Grove, C.S. Hopkinson, K.J. Nadelhoffer, D.E. Pataki, and D.P.C. Peters. 2008. The changing landscape: Ecosystem responses to urbanization and pollution across climatic and societal gradients. Frontiers in Ecology and the Environment 6(5):264-272. https://doi.org/10.1890/070147.
Hales, T.C., and C.F. Miniat. 2017. Soil moisture causes dynamic adjustments to root reinforcement that reduce slope stability. Earth Surface Processes and Landforms 42:803-813. https://doi.org/10.1002/esp.4039.
Hales, T.C., C.R. Ford, T. Hwang, J.M. Vose, and L.E. Band. 2009. Topographic and ecologic controls on root reinforcement. Journal of Geophysical Research 114: F03013. https://doi.org/10.1029/2008JF001168.
Healing, S., C.E. Benkwitt, R.E. Dunn, and N.A.J. Graham. 2024. Seabird-vectored pelagic nutrients integrated into temperate intertidal rocky shores. Frontiers in Marine Science 11. https://doi.org/10.3389/fmars.2024.1343966.
Heffernan, J.B., P.A. Soranno, M.J. Angilletta, Jr., L.B. Buckley, D.S. Gruner, T.H. Keitt, J.R. Kellner, et al. 2014. Macrosystems ecology: Understanding ecological patterns and processes at continental scales. Frontiers in Ecology and the Environment 12: 5-14. https://doi.org/10.1890/130017.
Holt, R.D. 1997. From metapopulation dynamics to community structure: Some consequences of spatial heterogeneity. Pp. 149-164 in Metapopulation Biology: Ecology, Genetics, and Evolution, I. Hanski and M.E. Gilpin, eds. San Diego: Academic Press.
Hou, W., T. Hu, L. Yang, X. Liu, X. Zheng, H. Pan, X. Zhang, S. Xiao, and S. Deng. 2023. Matching ecosystem services supply and demand in China’s urban agglomerations for multiple-scale management. Journal of Cleaner Production 420:138351. https://doi.org/10.1016/j.jclepro.2023.138351.
Hwang, T., L. Band, T.C. Hales, C.F. Miniat, J.M. Vose, P.V. Bolstad, B. Miles, and K. Price. 2015. Simulating vegetation controls on hurricane-induced shallow landslides with a distributed ecohydrological model. Journal of Geophysical Research: Biogeoscience 120:361-378.
Ibáñez, I., L. Petri, D.T. Barnett, E.M. Beaury, D.M. Blumenthal, J.D. Corbin, J. Diez, J.S. Dukes, R. Early, I.S. Pearse, C.J.B. Sorte, M. Vilà, and B. Bradley. 2023. Combining local, landscape, and regional geographies to assess plant community vulnerability to invasion impact. Ecological Applications 33(4):e2821. https://doi.org/10.1002/eap.2821.
Inamine, H., A. Miller, S. Roxburgh, A. Buckling, and K. Shea. 2022. Pulse and press disturbances have different effects on transient community dynamics. The American Naturalist 200:571-583. https://doi.org/10.1086/720618.
Jaffe, D.A., S.M. O’Neill, N.K. Larkin, A.L. Holder, D.L. Peterson, J.E. Halofsky, and A.G. Rappold. 2020. Wildfire and prescribed burning impacts on air quality in the United States. Journal of the Air & Waste Management Association 70: 583-615. https://doi.org/10.1080/10962247.2020.1749731.
Leibold, M.A., M. Holyoak, and N. Mouquet, P. Amarasekare, J.M. Chase, M. Hoopes, R. Holt, J. Shurin, D. Tilman, M. Loreau, and A. Gonzalez. 2004. The metacommunity concept: A framework for multi-scale community ecology. Ecology Letters 7:601-613. https://doi.org/10.1111/J.1461-0248.2004.00608.X.
Levin, S.A. 1992. The problem of pattern and scale in ecology: The Robert H. MacArthur Award Lecture. Ecology 73:1943-1967. https://doi.org/10.2307/1941447.
Levin, S. 2024. Ecological resilience. Encyclopedia Britannica. https://www.britannica.com/science/ecological-resilience (accessed September 3, 2024).
Liu, J. 2023. Leveraging the metacoupling framework for sustainability science and global sustainable development. National Science Review 10(7):nwad090. https://doi.org/10.1093/nsr/nwad090.
Liu, J., G. Daily, P. Ehrlich, and G.W. Luck. 2003. Effects of household dynamics on resource consumption and biodiversity. Nature 421:530-533. https://doi.org/10.1038/nature01359.
Liu, J., T. Dietz, S.R. Carpenter, M. Alberti, C. Folke, E. Moran, A.N. Pell, et al. 2007. Complexity of coupled human and natural systems. Science 317:1513-1516. https://doi.org/10.1126/science.1144004
Liu, J., V. Hull, W. Yang, A. Viña, X. Chen, Z. Ouyang, and H. Zhang (eds.). 2016. Pandas and People: Coupling Human and Natural Systems for Sustainability. Oxford University Press. https://doi.org/10.1093/acprof:oso/9780198703549.001.0001.
Locey, K.J., and J.T. Lennon. 2016. Scaling laws predict global microbial diversity. Proceedings of the National Academy of Sciences of the United States of America 113(21):5970-5975. https://doi.org/10.1073/pnas.1521291113.
Loreau, M., N. Mouquet, and R.D. Holt. 2003. Meta-ecosystems: A theoretical framework for a spatial ecosystem ecology. Ecology Letters 6:673-679. https://doi.org/10.1046/j.1461-0248.2003.00483.x.
MacDonald, G., T. Wall, C.A.F. Enquist, S.R. LeRoy, J.B. Bradford, D.D. Breshears, T. Brown, et al. 2023. Drivers of California’s changing wildfires: A state-of-the-knowledge synthesis. International Journal of Wildland Fire 32:1039-1058. https://doi.org/10.1071/WF22155.
Mann, M.E., Z. Zhang, M.K. Hughes, R.S. Bradley, S.K. Miller, S. Rutherford, and F. Ni 2008. Proxy-based reconstructions of hemispheric and global surface temperature variations over the past two millennia. Proceedings of the National Academy of Sciences of the United States of America 105(36):13252-13257. https://doi.org/10.1073/pnas.0805721105.
Martinez-Vilalta, J., and F. Lloret. 2016. Drought-induced vegetation shifts in terrestrial ecosystems: The key role of regeneration dynamics. Global and Planetary Change 144:94-108. https://doi.org/10.1016/j.gloplacha.2016.07.009.
Melillo, J.M., T.T. Richmond, and G. Yohe (eds.). 2014. Climate Change Impacts in the United States: The Third National Climate Assessment. U.S. Global Change Research Program. https://nca2014.globalchange.gov/report.
Miller, R.W. 2021. Thick smoke from western wildfires is traveling thousands of miles, clouding NYC skies. USA Today. https://www.usatoday.com/story/news/nation/2021/07/21/wildfire-smoke-cause-poor-air-quality-index-red-sun-nyc/8038354002/ (accessed March 8, 2024).
Myers, N., R. Mittermeier, C. Mittermeier, G.A.B. da Fonseca, and J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858. https://doi.org/10.1038/35002501.
NASEM (National Academies of Sciences, Engineering, and Medicine). 2023. Progress Toward Restoring the Everglades: The Ninth Biennial Review—2022. Washington, DC: The National Academies Press. https://doi.org/10.17226/26706.
Peters, D.P.C., P.M. Groffman, K.J. Nadelhoffer, N.B. Grimm, S.L. Collins, W.K. Michener, and M.A. Huston. 2008. Living in an increasingly connected world: A framework for continental-scale environmental science. Frontiers in Ecology and the Environment 6(5):229-237. https://doi.org/10.1890/070098.
Pickett, S.T.A., M.L. Cadenasso, and B. McGrath (eds). 2013. Resilience in Ecology and Urban Design: Linking Theory and Practice for Sustainable Cities. Dordrecht: Springer Science & Business Media.
Polasky, S., B. Bryant, P. Hawthorne, J. Johnson, B. Keeler, and D. Pennington. 2015. Inclusive wealth as a metric of sustainable development. Annual Review of Environment and Resources 40:445-466. https://doi.org/10.1146/annurev-environ-101813-013253.
Reid, R.S., M.E. Fernández-Giménez, and K.A. Galvin. 2014. Dynamics and resilience of range-lands and pastoral peoples around the globe. Annual Review of Environment and Resources 39:217-242. https://doi.org/10.1146/annurev-environ-020713-163329.
Remple, K.L., N.J. Silbiger, Z.A. Quinlan, M.D. Fox, L.W. Kelly, M.J. Donahue, and C.E. Nelson. 2021. Coral reef biofilm bacterial diversity and successional trajectories are structured by reef benthic organisms and shift under chronic nutrient enrichment. NPJ Biofilms Microbiomes 7:84. https://doi.org/10.1038/s41522-021-00252-1.
Rose, K.C., R.A. Graves, W.D. Hansen, B.J. Harvey, J. Qui, S.A. Wood, C. Ziter, M.G. Turner. 2016. Historical foundations and future directions in macrosystems ecology. Ecology Letters 20(2):147-157. https://doi.org/10.1111/ele.12717.
Seto, K.C., B. Güneralp, and L.R. Hutyra. 2012. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proceedings of the National Academy of Sciences of the United States of America 109:16083-16088. https://doi.org/10.1073/pnas.1211658109.
Shaffer, J.P., L.-F. Nothias, L.R. Thompson, et al. 2022. The Earth Microbiome Project 500 Consortium. Standardized multi-omics of Earth’s microbiomes reveals microbial and metabolite diversity. Nature Microbiology 7:2128-2150. https://doi.org/10.1038/s41564-022-01266-x.
Shepherd, J.M., H. Pierce, and A.J. Negri. 2002. Rainfall modification by major urban areas: Observations from spaceborne rain radar on the TRMM satellite. Journal of Applied Meteorology and Climatology 41:689-670. https://doi.org/10.1175/1520-0450(2002)041%3C0689:RMBMUA%3E2.0.CO;2.
Stallard, R.F. 1998. Terrestrial sedimentation and the carbon cycle: Coupling weathering and erosion to carbon burial. Global Biogeochemical Cycles 12(2):231-257. https://doi.org/10.1029/98GB00741.
Tavárez, H., L. Elbakidze, O.J. Abelleira-Martínez, Z. Ramos-Bendaña, and N.A. Bosque-Pérez. 2022. Willingness to pay for gray and green interventions to augment water supply: A case study in rural Costa Rica. Environmental Management 69:636-651. https://doi.org/10.1007/s00267-021-01476-9.
Thompson, L.R., J.G. Sanders, D. McDonald, et al. 2017. The Earth Microbiome Project Consortium. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 551:457-463. https://doi.org/10.1038/nature24621.
Turner, B.L., R.E. Kasperson, P.A. Matson, J.J. McCarthy, R.W. Corell, L. Christensen, N. Eckley, et al. 2003. A framework for vulnerability analysis in sustainability science. Proceedings of the National Academy of Sciences of the United States of America 100:8074-8079. https://doi.org/10.1073/pnas.1231335100.
Turner, B.L., E.F. Lambin, and A. Reenberg. 2007. The emergence of land change science for global environmental change and sustainability. Proceedings of the National Academy of Sciences of the United States of America 104:20666-20671. https://doi.org/10.1073/pnas.0704119104.
Turner, R.K., and G.C. Daily. 2008. The ecosystem services framework and natural capital conservation. Environmental and Resource Economics 39:25-35. https://doi.org/10.1007/s10640-007-9176-6.
UNEP (United Nations Environment Programme). 2018. Inclusive Wealth Report: Measuring Sustainability and Well Being. Nairobi, Kenya.
Viña, A., M.-N. Tuanmu, W. Xu, Y. Li, Z. Ouyang, R. DeFries, and J. Liu. 2010. Range-wide analysis of wildlife habitat: Implications for conservation. Biological Conservation 143:1960-1969. https://doi.org/10.1016/j.biocon.2010.04.046.
Vitousek, P.M. 2004. Nutrient Cycling and Limitation. Hawai’i as a Model System. Princeton, N.J.; Princeton University Press.
Wang, C., Y. Ye, and Z. Huang. 2024. Synergistic development in the Guangdong-Hong Kong-Macao Greater Bay Area: Index measurement and systematic evaluation based on industry-innovation-infrastructure-institution perspectives. Journal of Cleaner Production 434: 140093. https://doi.org/10.1016/j.jclepro.2023.140093.
Xiao, H., S. Bao, J. Ren, Z. Xu, S. Xue, and J. Liu. 2024. Global transboundary synergies and trade-offs among Sustainable Development Goals from an integrated sustainability perspective. Nature Communications 15:500. https://doi.org/10.1038/s41467-023-44679-w.
Xu, W., A. Viña, L. Kong, S.L. Pimm, J. Zhang, W. Yang, Y. Xiao, L. Zhang, X. Chen, J. Liu, and Z. Ouyang. 2017. Reassessing the conservation status of the giant panda using remote sensing. Nature Ecology & Evolution 1:1635-1638. https://doi.org/10.1038/s41559-017-0317-1.
Xu, Z., Y. Li, S.N. Chau, T. Dietz, C. Li, L. Wan, J. Zhang, L. Zhang, Y. Li, M.G. Chung, and J. Liu. 2020. Impacts of international trade on global sustainable development. Nature Sustainability 3:964-971. https://doi.org/10.1038/s41893-020-0572-z.
Yang, W., T. Dietz, W. Liu, J. Luo, and J. Liu. 2013. Going beyond the Millennium Ecosystem Assessment: An index system of human dependence on ecosystem services. PLoS ONE 8(5):e64581. https://doi.org/10.1371/journal.pone.0064581.
Yang, H., A. Viña, J.A. Winkler, M.G. Chung, Y. Dou, F. Wang, J. Zhang, Y. Tang, T. Connor, Z. Zhao, and J. Liu. 2019. Effectiveness of China’s protected areas in reducing deforestation. Environmental Science and Pollution Research 26:18651-18661. https://doi.org/10.1007/s11356-019-05232-9.
Yang, H., A. Viña, J.A. Winkler, M. G. Chung, Q. Huang, Y. Dou, W.J. McShea, M. Songer, J. Zhang, and J. Liu. 2021. A global assessment of the impact of individual protected areas on preventing forest loss. Science of the Total Environment 777:145995. https://doi.org/10.1016/j.scitotenv.2021.145995.
Zhang, J., V. Hull, J. Huang, W. Yang, S. Zhou, W. Xu, Y. Huang, Z. Ouyang, H. Zhang, and J. Liu. 2014. Natural recovery and restoration in giant panda habitat after the Wenchuan earthquake. Forest Ecology and Management 319:1-9. https://doi.org/10.1016/j.foreco.2014.01.029.
Zhang, J., V. Hull, J. Huang, S. Zhou, W. Xu, H. Yang, W.J. McConnell, R. Li, D. Liu, Y. Huang, Z. Ouyang, H. Zhang, and J. Liu. 2015 Activity patterns of the giant panda (Ailuropoda melanoleuca). Journal of Mammalogy 96(6):1116-1127. http://dx.doi.org/10.1093/jmammal/gyv118.
Zhao, Z., M. Cai, F. Wang, J.A. Winkler, T. Connor, M.G. Chung, J. Zhang, H. Yang, Z. Xu, Y. Tang, Z. Ouyang, H. Zhang, and J. Liu. 2021. Synergies and tradeoffs among Sustainable Development Goals across boundaries in a metacoupled world. Science of the Total Environment 751:141749. https://doi.org/10.1016/j.scitotenv.2020.141749.

























{kind=link}