Out of Thin Air: Dinosaurs, Birds, and Earth's Ancient Atmosphere (2006)
Chapter: 8 The Triassic Explosion
8
THE TRIASSIC EXPLOSION
The animals and plants of the Triassic Period make up a most interesting assemblage of organisms. This chapter looks at this crossroad in time. Coming out of the most devastating of all mass extinctions, the early Triassic world was empty of life. At the same time, all modeling suggests that a long interval of the Triassic was a time when oxygen levels were lower than today. In Chapter 2 it was proposed that times of low oxygen, especially following mass extinction, foster disparity: the diversity of new body plans. In the Triassic these two factors combined to create the largest number of new body plans seen since the Cambrian. This chapter proposes that it is to that seminal Cambrian time that we can most accurately compare the Triassic. We should give this time and its biotic consequences a new name.
THE TRIASSIC EXPLOSION
The middle Triassic was a time of amazing disparity on land and in the sea. In the latter, new stocks of bivalved mollusks took the place of the many extinct brachiopods, while a great diversification of ammonoids and nautiloids refilled the oceans with active predators. Fully a quarter of all the ammonites that ever lived have been found in Triassic rocks—a time interval that is only 10 percent of their total time of existence on Earth. The oceans filled with their kind, in shapes and
patterns completely new compared to their Paleozoic ancestors—and why not? As shown above, this kind of animal was the preeminent, low-oxygen adaptation among all invertebrates. A new kind of coral, the scleractinians, began to build reefs, and many land reptiles returned to the sea. But it is on land that the most sweeping changes in terms of body plan replacements—and body plan experimentation—took place. Never before and never since has the world seen such a diverse group of different anatomies on land. Some were familiar Permian types: the therapsids that survived the Permian extinction diversified and competed with archosaurs for dominance of the land early in the Triassic, but this ascendance was short lived. Many kinds of reptiles were locked in a competitive struggle with them and with each other for land dominance. From mammal-like reptiles to lizards, earliest mammals to true dinosaurs, the Triassic was a huge experiment in animal design.
Why was this? The conventional answer is that the Permian extinction removed so many of the dominant land animals that it opened the way for more innovation than at any other nonextinction time, perhaps any other mass extinction time as well. It was the most devastating of the mass extinctions. Perhaps, as well, it was simply that many terrestrial animal body plans finally came to an evolutionary point of really working efficiently rather than the sprawling posture of the early reptiles and amphibians. Even as late as the end of the Permian and into the Triassic, groups as mature as the dicynodonts and cynodonts were still trying to attain the most efficient kind of upright posture—rather than the less efficient, splayed-leg orientation of the land reptiles—with all of the ramifications and penalties in respiration that this entailed. But perhaps there is more than this. Body plans were being stimulated into creation by intense selective pressures, and dominant among these was the need to access sufficient oxygen to feed, breed, and compete in a low-oxygen world. There is an old adage about nothing sharpening the mind faster than imminent death. The same might be said about evolutionary forces when faced with the most pressing of all selective pressures—attaining the oxygen necessary for the high levels of animal activity that had been evolutionarily attained in the high-oxygen world of the Permian, when nothing was
easier to extract from the atmosphere. The two-thirds drop in atmospheric oxygen certainly lit the fuse to an evolutionary bomb, which exploded in the Triassic.
Thus, the diversity of Triassic animal plans is analogous to the diversity of marine body plans that resulted from the Cambrian Explosion. It also occurred for nearly the same reasons and, as will be shown, was as important for animal life on land as the Cambrian Explosion was for marine animal life. As we saw in Chapter 3, the Cambrian Explosion followed a mass extinction (of the Ediacaran fauna), and it was a time of lower oxygen than today. The latter stimulated much new design. Finally, the Cambrian Period itself ended in a mass extinction—mainly of trilobites that we know of but also among many of the more exotic arthropods known from the Burgess Shale, such as Anomalocaris. In similar fashion, on land the Triassic Explosion followed a mass extinction, was a time of lower oxygen, and ended in a mass extinction. Before this extinction, mammals had evolved, as had true dinosaurs, but many of the other kinds of body plans disappeared and dinosaurs, were the dominant land animals. In this end-Triassic mass extinction the dinosaurs suffered least of all. Why dinosaurs? This chapter will look at those questions.
VOYAGE
Let’s begin by looking back to the middle part of the Triassic period. In this middle-late Triassic world, 215 million years ago, on land at least we seem to have arrived among a veritable smorgasbord of animal body plans. Many quite different kinds of vertebrates inhabit this world. Dog-like creatures walk beneath the conifer- and tree-fern dominated vegetation. They are cynodonts, carnivorous varieties, but there are massive herbivores belonging to the same group as well. They are all very mammalian in appearance and behavior, except in one aspect. They move little and seem to tire easily. The carnivores mostly lay in wait, and the herbivores browse stolidly. The cynodonts are not the only mammal-reptiles here, for rhino-sized dicynodonts also browse the low brushy vegetation. Their odd, name-giving tusks extending from a parrot-like beaked mouth make them look like nothing of our
world, and seeing them harkens memories of the late Permian world prior to the great Permian mass extinction. All are panting heavily and give the impression of animals having just engaged in strenuous exercise. But most have been motionless; yet they pant for good reason. The level of atmospheric oxygen at this time is equivalent to being higher than 10,000 feet in our world. Except that here we are not atop any mountain: the low swamps and nearby arm of the sea attest to our being at sea level.
Other herbivores are here too, and they are clearly from groups long-diverged away from the mammal-like reptile lineage. It is soon apparent that they are more numerous than the mammal-like reptiles. One of the oddest is another beaked herbivore, from the reptile group known as rhynchosaurs, and near it is a heavily armored quadruped, an aetosaur, looking something like an armadillo, only much larger and better armored. Soon we notice other quadruped reptiles, all fairly primitive archosaurs. Many are 5 to 10 feet in length, and they too move little; when they do, the movement is labored and the panting rapid. Large size and armor evolve for one reason, to avoid being eaten—but the cost is high. Moving a heavy body about extracts a great metabolic cost. Yet there is method in this seemingly morphological madness, for it is clear that the carnivores here are in abundance.
A number of reptiles are visible with heads like that of a crocodile but with bodies obviously evolved for rapid movement on land. Some rise up on their back legs, but they are still quadrupeds for rapid movement. They seem better suited for prolonged movement than the other designs seen till until now, but they are no greyhounds. They prowl but in labored fashion.
Until now all the animals we have seen have been quadrupeds, but it is not long before we encounter our first bipedal animal, and soon we find that this world certainly has its share of animals that walk bipedally. All of them seem far more at ease in this atmosphere, one that bothers even us humans, supposedly advanced mammals that we are. We have finally encountered our first dinosaurs.
Some are small and carnivorous; others, the prosauropods, are relative giants, the largest animals on land and the largest animals ever evolved up to this point. There seem to be many varieties of the smaller
bipedal forms, and we are thankful that most of the obvious carnivores are relatively small, for they zip about, running rings around the other vertebrates. But soon a much larger form, 12 feet in length, strides into view, a staurikosaur, the top carnivore among the dinosaurs. We watch this animal, puzzled at how different it seems from the nondinosaurs. Despite its size, it is very active.
This dichotomy extends as well into the swampy and freshwater habitats, where untold numbers of crocodile-like phytosaurs loll on the banks. They too seem not disposed to frolic in any fashion. It is only the dinosaurs that move about with speed, grace, and purpose.
In the oceans we see a major change. Largely gone are the brachiopods, the bivalved invertebrates so dominant in Paleozoic oceans. In their place is another kind of bivalve—the more familiar clams. Few burrow. Most rest on the surface of the sea bottom, sometimes in huge numbers. Swimming above the bottom are many varieties of fish, among them a host of ammonites. This latter group just missed total extinction in the Permian extinction. From the few spared species, however, a host of new species has evolved until now. In the latter parts of the Triassic period, they are even more numerous than anytime before.
There are also coral reefs again, but like the bottom communities of invertebrates found on the sandier and muddier bottoms, the reefs are composed of an entirely new suite of corals. Gone are the tabulate

Reconstruction of a ceratite ammonite, agroup found only during the Triassic.These forms evolved from the few ammonites that survived the Permian extinction, an event that decimated thecephalopods.
and horn corals, they are replaced by the scleractinians, forms that will persist to our time.
As interesting as the invertebrates and even the fish are of this world, it is the larger vertebrates in the sea that really give pause. Diverse reptilian stocks have obviously returned to the ancestral ocean. Some, the stocky placodonts, resemble large, clumsy seals as they swim down and root through the clam beds with their peg-like teeth. But the stars of the show are the ichthyosaurs, reptiles that have so thoroughly evolved for a swimming habit that they have lost their legs entirely and are the most fish-like of originally terrestrial vertebrates that the world has ever known or will know until the eventual evolution of dolphins, far, far into the future from this Triassic time. While many are small, there are larger forms too: Mixosaurus is 30 feet long but it is dwarfed by the monstrous Shonisaurus, some of which reach a length of 60 feet. This huge ichthyosaur rivals sperm whales for the title of world’s largest aquatic predator of all time, and it preys at will on hosts of fish and smaller reptiles. It has dinner-plate-sized eyes, the largest eyes ever evolved either before or after this.
The marine world, at least, despite its strange and frightful beasts, strikes a sense of some familiarity. But even here we begin to notice strange behavior: the diving reptiles, like the placodonts, come to the surface frequently to breath, as do even the ichthyosaurs. The low oxygen takes its toll.
TRIASSIC REBOUND
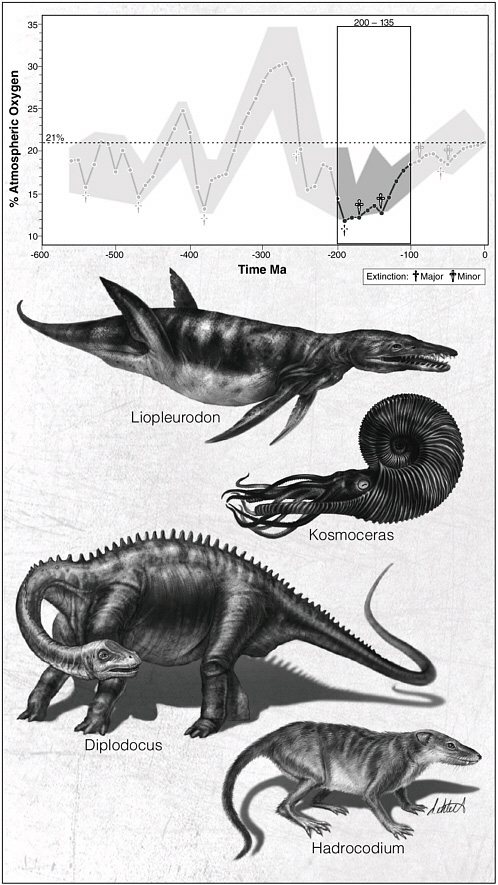
The oxygen story for the Triassic is stunning. Oxygen dropped to minimal levels of between 10 and 15 percent and then stayed there for at least 5 million years, from 245 million to 240 million years ago.
The officially designated early Triassic time interval was from 250 million to about 245 million years ago. During this time there was little in the way of recovery from the Permian extinction. There is also a very curious record of large-scale oscillations in carbon isotopes from this time, indicating that the carbon cycle was being perturbed in what looks like either methane gas entering the oceans, or a succession of small-scale extinctions taking place. All evidence certainly paints a picture of a stark and environmentally challenging world for animal life.
Microbes may have thrived, especially those that fixed sulfur, but animals had a long period of difficult times. But difficult times are what best drive the engines of evolution and innovation and from this trough in oxygen on Earth emerged new kinds of animals, most sporting respiratory systems better able to cope with the extended oxygen crisis. On land two new groups were to emerge from the wreckage: mammals and dinosaurs. The former would become bridesmaids in waiting while the latter would take over the world.
THE DIFFERENT FATES OF THETRIASSIC THERAPSIDS AND DIAPSIDS
As we saw in Chapter 7, the Permian extinction nearly annihilated land life. The therapsids were hit hard. Much less is known about the diapsids, for at the end of the Permian they were a rare and little seen group in the areas, such as the Karoo or Russia, that have yielded rich deposits with abundant dicynodont (therapsid) faunas. In the Karoo at least, only small fragments of diapsids have come from our uppermost Permian study sections, although two skulls now being prepared as I write this may turn out to be forms that would give rise to the great diapsids and dinosaur dynasty. Roger Smith found them in highest Permian rocks on our last joint collecting trip from the same section where the lowest Triassic diapsids was found. Are they the same species? We will soon know.
If we are still poorly informed about their Permian ancestry, there is no ambiguity about the success of the earliest Triassic diapsids. In the Karoo, in strata only a few meters above the beds that seem to mark the transition from Permian to Triassic, there are relatively common remains of a fairly large reptile known as Proterosuchus (also known as Chasmatosaurus). This was definitely a land animal with a very impressive set of sharply pointed teeth. It was also definitely a predator, but like a crocodile, its legs were splayed to the sides (if somewhat more upright than the crocodilian condition). But this condition was to rapidly change in the diapsids to a more upright orientation as the Triassic progressed, and more gracile and rapid predators soon replaced the early diapsids such as Proterosuchus.
While the need for speed was surely a driver toward this better
locomotory posture, just as important may have been the need to be able to breathe while walking. Like a lizard, Proterosuchus may still have had a back-and-forth sway to its body as it walked, and this sort of locomotion causes compression on the lung area due to what is known as Carrier’s Constraint—the concept that quadrupeds with splayed out legs, such as most (but not all!) lizards, cannot breathe while they run, because their sinuous, side-to-side swaying impinges on the lungs and rib cage, inhibiting inspiration.
For this reason, most lizards and salamanders cannot breathe while walking, and Proterosuchus may have had something of this effect, although not as pronounced as in modern-day salamanders or lizards. A solution is to put the legs beneath, but this is only a partial solution. To truly be free of the constraint that breathing puts on posture, extensive modifications to the respiratory system and the locomotory system had to be made. The lineage that led to dinosaurs and birds found an effective and novel adaptation to overcome this breathing problem: bipedalism. By removing the quadruped stance, they were freed of the constraints of motion and lung function. The ancestors of the mammals also made new innovations, including a secondary palate (which allows simultaneous eating and breathing) and a complete upright (but still quadruped) stance. But this was still not satisfactory and a new kind of breathing system was evolved. A powerful set of muscles, known as the diaphragm, allowed a much more forceful system for inspiring and then exhaling air.
Thus, by the middle Triassic, some very different respiratory designs were in play, with natural selection and competition as the arbiters. We know what kind of lungs the mammals had. But what about the diapsids—and their most famous members (and descendents of the early kinds such as Proterosuchus)—the dinosaurs? By the end of the middle Triassic they had burst upon the scene. How did they breathe? Therein lies a controversy.
What kind of lungs did the earliest dinosaurs evolve? What changes to this lung design came about in their descendents? This has been the source of controversy for more than two decades now. But before we enter the debate, it should be noted that whichever lung system was found in the Triassic dinosaurs, it evolved for a self-similar
reason: oxygen levels reached their lowest point in the Triassic, coincidentally at the time when a majority of Permian land animals were going extinct (leaving many empty niches and thus wholesale evolutionary and ecological opportunity), and many vertebrates responded to these two factors by rapidly producing a host of new kinds of vertebrate body plans—and respiration systems as well. The most famous of the new Triassic body plans was a bipedal form that we call dinosaurs.
WHAT IS A DINOSAUR?
Because of its general interest and rather sensational aspects, perhaps the most commonly asked question about dinosaurs is the manner of their extinction. The 1980 hypotheses by the Alvarez group that Earth was hit 65 million years ago by an asteroid and that the environmental effects of that asteroid rather suddenly caused the Cretaceous-Tertiary mass extinction in which the dinosaurs were the most prominent victims, keeps this question paramount in people’s minds. The fact that this controversy is rekindled every several years by some new finding brings it to the surface once again. Thus its preeminence even supercedes the question of whether or not the dinosaurs were warm-blooded. Way down on the list of questions about dinosaurs is the inverse of the extinction question—not why they died out, but why they evolved in the first place.
We know when they first appeared, in the second third of the Triassic Period (some 235 million years ago), and we know what these earliest dinosaurs looked like: most were like smaller versions of the later and iconic Tyrannosaurus rex and Allosaurus. They were bipedal forms that quickly became large. What has not been largely known or even considered is the new understanding that 230 million years ago was the time when oxygen may have been nearing its lowest level since the Cambrian Period.
So here is a new view here: dinosaurs evolved during, or immediately before, the Triassic oxygen low, a time when oxygen was at its lowest value of the last 500 million years—and their body plan is a result of adaptation to low oxygen.
Many other animals changed body plans in response to extremes in oxygen and so too did the dinosaurs, in my view. The dinosaur body plan is radically different from earlier reptilian body plans and appears in virtually a dead heat (and in great global heat) with the oxygen minimum. Perhaps this is a coincidence. But because many of the aspects of “dinosaurness” can be explained in terms of adaptations for life in low oxygen, that seems unlikely.
To formalize this: the initial dinosaur body plan (evolved first by saurischian dinosaurs such as Staurikosaurus and the somewhat younger Herrerasaurus) was in some part in response to the low-oxygen conditions of the time:
Hypothesis 8.1: The initial dinosaur body plan of bipedalismevolved as a response to low oxygen in the middle Triassic. Witha bipedal stance the first dinosaurs overcame the respiratorylimitations imposed by Carrier’s Constraint. The Triassic oxygen low thus triggered the origin of dinosaurs through the formation of this new body plan.
The fossil record shows that the earliest true dinosaurs were bipedal and came from more primitive bipedal thecodonts slightly earlier in the Triassic. These thecodonts (diapsids) were the ancestors of the lineage giving rise to the crocodiles as well and may have been either warm-blooded or heading that way. Bipedalism was a recurring body plan in this group, and there were even bipedal crocodiles early on. Why bipedalism, and how could it have been an adaptation to low oxygen?
Earlier we saw how even most modern-day lizards cannot breath while they run, and this is due to their sprawling gait. Modern-day mammals show a distinct rhythm by synchronizing breathtaking with limb movement. Horses, jackrabbits, and cheetahs (among many other mammals) take one breath per stride. Their limbs are located directly beneath the mass of the body and to allow this the backbone in these quadruped mammals has been enormously stiffened compared to the backbones of the sprawling reptiles. The mammalian backbone bows slightly downward and then straightens out with running, and this slight up-and-down bowing is coordinated with air inspiration and
exhalation. But this system did not appear until true mammals appeared in the Triassic. Even the most advanced cynodonts of the Triassic were not yet fully upright and thus would have suffered somewhat when trying to run and breathe.
By running on two legs instead of four, the lungs and rib cage are not affected. Breathing can be disassociated from locomotion. The bipeds can take as many breaths as they need to in a high-speed chase. At a time of low oxygen but high predation, any slight advantage either in chasing down prey, or in running from predators—even in the amount of time looking for food, or how food is looked for, would surely have increased survival. The sprawling predators of the late Permian, such as the fearsome gorgonopsians, were, like most predators during and before their time, ambush predators, as all lizards are today. So what must it have been like for the animals of the Triassic when they found that, for the first time, the predators were out searching for them rather than hiding and waiting? Were any smart enough to register surprise? The results: carnage, surely carnage. This is why dinosaurs may have arisen.
All dinosaurs descended from bipedal ancestors, even the massive quadrupeds of later in the Mesozoic. In the Triassic, the crocodile lineage and the dinosaur lineage shared a quadruped common ancestor. This beast may have been a reptile from South Africa named Euparkeria. This group is technically called the Ornithodira, and even the earliest members began to evolve toward bipedalism. This is shown by their ankle bones, which simplified into a simple hinge joint from the more complex system found in quadrupeds. This, accompanied by a lengthening of the hind limbs relative to the forelimbs, is also evidence of this trend, as is the neck, which elongates and forms a slight S shape. These early Ornithodira themselves split into two distinct lineages. One took to the air. These were the pterosaurs, and the late Triassic Ornithodira named Scleromochlus might have been the very first of its kind, a still-terrestrial form that looks like a fast runner that perhaps began gliding between long steps using arms with skin flaps. The oldest undoubtedly flying pterosaur was Eudimorphodon, also of the late Triassic.
While these ornithodires edged toward flight, their terrestrial sis-
ter group headed toward the first dinosaur morphology. The Triassic Lagosuchus was a transitional form between a bipedal runner and a quadruped. It probably moved slowly on all fours but reared up on its hind legs for bursts of speed—the bursts necessary to bring down prey, for this was a predator. But it still had forelimbs and hands that had not yet attained the dinosaur type of morphology, so it is not classified as a dinosaur. Its successor, the Triassic Herrerasaurus, meets all the requirements and is classified as a dinosaur—the first. As we shall see below, it may have lacked one attribute that its immediate descendents would rectify: a new kind of respiratory system that could handle the still-lowering oxygen content of Earth’s atmosphere.
This first dinosaur was fully bipedal, and it could grasp objects with its hands, since it had a thumb like we do. The evolution of this five-fingered hand, being very distinct from the functionally three-toed foot (there were five actual toes, but two were so vestigial that only three toes touched the ground while running or walking), was a new innovation. Because it was not totally bipedal, evolution no longer had to worry about maintaining a hand that had to touch the ground for locomotion. So with a free appendage no longer necessary for locomotion, what to do with it? The much later and more famous T. rex reduced the size of the forearm to the point that some have suggested that it was non-functional. Not so for these first dinosaurs, however. While their posture was that of the later carnivorous dinosaurs so familiar to us, their hands were obviously used—probably for catching and holding prey while on the run.
So this is the body plan of the first dinosaurs, from which all the rest evolved: bipedal, elongated neck, grasping hands with a functioning thumb, a large and distinctive pelvis for the massive muscles, and a large surface area needed for those muscles used in walking and running. These early bipeds were relatively small, and before the end of the Triassic they again split into two groups, which remained the most fundamental split of the entire dinosaur clan. A species of these bipedal Triassic dinosaurs modified its hip bones to incorporate a backturned pubis, compared to the forward-facing pubis of the first dinosaurs. As any schoolboy knows, this change in pelvic structure marks the division of the dinosaurs into the two great divisions: the ancestral
saurischians and their derived descendents, with whom they would share the world for about the next 170 million years, the ornithischians.
Of interest here, of course, is how dinosaurs breathed. First, we must look at vertebrate lungs and then tackle the controversy over dinosaur lungs.
AMNIOTE LUNGS
The lungs of modern-day amniotes (reptiles, birds, and mammals) are of two basic types (although we will see that there are more than two respiratory systems, which include lungs, circulatory system, and blood pigment type). Both kinds of lungs can be reasonably derived from a single kind of Carboniferous reptilian ancestor that had simple sac-like lungs. Extant mammals all have alveolar lungs, whereas extant turtles, lizards, birds, and crocodiles—all the rest—have septate lungs.
Alveolar Lungs These lungs consist of millions of highly vascularized, spherical sacs called alveoli. Air flows in and out of the sacs; it is therefore bidirectional, a characteristic of alveolar lungs. Mammals use this system, and our familiar breathing in, out, in, out is quite typical. Nothing extraordinary here about us humans (compared to most other mammals, that is). The trick, of course, is that air must be pulled into these sacs and then expelled again as oxygen switches place with carbon dioxide. We do this by a combination of rib cage expansion (powered by muscles, of course) and contraction of the large suite of muscles collectively called the diaphragm. Somewhat paradoxically, contraction of the diaphragm causes the volume of the lungs to increase. These two activities, the interacting rib expansion and diaphragm contraction create a reduction in air pressure in the lung volume and air flows in. Exhaling is partially accomplished by elastic rebound of the individual alveoli. When they inflate, they enlarge, and soon after they naturally contract due to the elastic properties of their tissue. The many alveoli used in this kind of lung allow for a very efficient oxygen acquisition system, which we warm-blooded mammals very much need in order to maintain our active, movement-rich life styles.
Septate Lungs In contrast to the mammalian lung, the septate lung found in reptiles and birds is like one giant alveolus. To break it into
smaller pockets that increase surface area for respiratory exchange, a large number of blade-like sheets of tissue extend into the sac. These partitioning elements are the septa, which give these kinds of lungs their name. There are many variations on this basic lung design among the many different kinds of animals that use it. Some kinds of septate lungs are partitioned into small chambers; others have secondary sacs that rest outside the lung but are connected to it by tubes. As in the alveolar lung, airflow is bidirectional but a difference is that the septate lungs are not elastic and thus do not naturally contract in size following inhalation. Lung ventilation also varies across groups with the septate lung. Lizards and snakes use rib movement to draw air in, but as we have seen, locomotion in lizards inhibits complete expansion of the lung cavity, and thus lizards do not breathe while moving.
The variety of modifications of the septate lung makes this system more diverse than the alveolar system. For instance, crocodiles have both a septate lung and a diaphragm—an organ not found in snakes, lizards, or birds. But the crocodile diaphragm is also somewhat different from that in mammals. It is not muscular but is attached to the liver and movement of this liver/diaphragm acts like a piston to inflate the lungs, with muscles attaching to the pelvis. (The mammalian [including human] diaphragm pulls the liver in the same way a crocodilian one does, creating a visceral piston, but the way this is accomplished differs in crocodiles and mammals.)
The Avian Air Sac System The last kind of lung found in terrestrial vertebrates is a variant on the septate lung. The best example of this kind of lung, and its associated respiratory system, is found in all birds. In this system the lungs themselves are small and somewhat rigid. Thus, bird lungs do not greatly expand and contract as ours do on each breath. But the rib cage is very much involved in respiration and especially those ribs closest to the pelvic region are very mobile in their connection to the bottom of the sternum, and this mobility is quite important in allowing respiration. But these are not the biggest differences. Very much unlike extant reptiles and mammals, these lungs have appendages added known as air sacs, and the resultant system of respiration is highly efficient. Here is why.
We mammals (and all other nonavians) bring air into our dead-end lungs and then exhale it. Birds have a very different system. When
a bird inspires air, it goes first into the series of air sacs. It then passes into the lung tissue proper, but in so doing the air passes but one way over the lung, since it is not coming down a trachea but from the attached air sacs. Exhaled air then passes out of the lungs. The one-way flow of air across the lung membranes allows a countercurrent system to be set up—the air passes one direction, and blood in the blood vessels in the lungs passes in the opposite direction. This countercurrent exchange allows for more efficient oxygen extraction and carbon dioxide venting than are possible in dead-end lungs. The air sacs are not involved in removing oxygen; they are an adaptation that allows the countercurrent system to work.
There is no question that the greater efficiency of this system (compared to all other lungs in vertebrates) is related to the two-cycle, countercurrent system produced by the air sac-lung anatomy in birds. But when did this system first appear and in how many groups? Therein lies the controversy.
The bones that in birds house air sacs (or parts thereof) should provide fossil evidence. But does a hole in a bone mean there were air sacs? Some of the air sacs, such as the abdominal air sacs in modern birds, leave no record of themselves in hard parts and thus could have been present in extinct groups without leaving a trace. So what was the situation in dinosaurs? There are two different camps: one proposes that dinosaurs had air sacs (Bakker and Paul, among others), the other that the dinosaurs had a simpler reptilian lung system (John Ruben and others).
When did this superb adaptation for low oxygen first evolve? This question is at the crux of a very contentious dispute between three scientists who have taken two very different positions about the kind of lungs that dinosaurs may have had. On one side is John Ruben of Oregon State University, an expert on reptile physiology and respiration, who contends that the flow through avian lung with its many auxiliary air sacs did not appear until the Cretaceous, some 100 million years ago—and then was found only in birds of that time. On the other side of this dispute are two workers profoundly interested in dinosaurs—Robert Bakker, author of The Dinosaur Heresies and champion of warm-blooded dinosaurs, and Gregory S. Paul, noted artist
and student of predatory dinosaurs, whose book Dinosaurs of the Air explicitly looks at this problem.
There is no love lost between these two sides, and the controversy has spilled over into the many Internet dinosaur posts. But an interesting new angle to this debate is that neither side entered into the argument by considering that oxygen levels may have fluctuated in the past, or even that the different kinds of lungs would favor or inhibit various kinds of animals living at different altitudes. In fact, it appears that both sides implicitly believe that oxygen levels in the early to middle Mesozoic may have been higher than now. This topic is very pertinent to our discussion here.
AIR SACS IN DINOSAURS?
As we saw above, the respiratory system of modern birds is composed of small lungs that have appendages—air sacs, which are also used for respiration. Together, the lungs and air sacs extract more oxygen than do the lungs of any other land animal. It has been estimated that at sea level a bird is 33 percent more efficient at extracting oxygen from air than a mammal. At higher altitudes the differential increases. At 5,000 feet, a bird may be 200 percent more efficient at extracting oxygen than a mammal. This gives birds a huge advantage over mammals at living at altitude. If such a system were present in the deep past, when oxygen levels even at sea level were lower than are found today at 5,000 feet, surely such a design would have been advantageous, perhaps enormously so, to a group that had it in competing or preying on groups that did not.
We know that birds evolved from small bipedal dinosaurs that were of the same lineage as the earliest dinosaurs—a group called saurischians. The first bird skeletons are from the Jurassic. But the air sacs attached to bird lungs are soft tissue and would fossilize only under the most unusual circumstances of preservation. Thus, we do not have direct evidence for when the air sac system came about. But we do have indirect evidence, enough to have stimulated the air sac in dinosaurs group to posit that all saurischian dinosaurs had the same air sac system, as do modern birds. And, like birds, they were warm-blooded. The evidence comes from holes in bones, places where these air sacs
may have rested. The Ruben group, however, vehemently opposes this view, suggesting, instead, that the air sac evolved only in the Cretaceous. A very distinct line in the sand was drawn, and the resulting controversy became juicy and nasty and, as will be shown below, is now resolved. Let’s look at this controversy in detail, as it impinges on many of the themes of this book so far.
The avian respiratory system with its various air sacs has been called an air sac system, and subsequently we will refer to it as such. The debate about when it first evolved began in the early 1970s with a young and energetic Robert Bakker, who made the breathtaking suggestion that some dinosaurs had the air sac system too. His evidence was as follows.
It had been known since the late 1800s that some dinosaur bones had curious hollows in them—just as bird bones do. For decades this discovery was either forgotten or attributed to an adaptation for lightening the massive bones, for many of these bones with holes—later called pneumatic bones—came from the largest land animals of all time, the giant sauropods of the Jurassic and Cretaceous. The pneumatic bones were found mainly in vertebrae. Birds have similar pneumatic vertebrae, and while it can be said that some bird bones were light to enhance flying, it was also clear that some of the air sacs attached to bird lungs rested in hollows in bones. Thus, in birds, bone pneumaticity was an adaptation for stashing away the otherwise space-taking air sacs. The bodies of animals are filled with necessary organs and putting the air sacs in hollowed-out bones made a lot of evolutionary sense. But Bakker made the leap and suggested that the pneumatic bones in his beloved fossil sauropods had evolved for a similar purpose and were direct evidence that sauropods had and used the air sac system. He was quite explicit, writing (in his stimulating 1986 book, The Dinosaur Heresies):
The dinosaur’s vertebral hollows are so similar to birds’ that there can be little doubt an avian style system of air sacs was at work in these Mesozoic animals. Moreover, the holes in bones represent the periphery of the total system. Birds locate their largest air sacs between their flight muscles and in their body cavity. Some dinosaurs (duckbills, horned dinosaurs) exhibited no vertebrate body but I suspect they had located air sacs fully within their body cavity.
Bakker thus put not only those dinosaurs with pneumatized bones (the group known as saurischians) into the air sac contingent but also the other major dinosaur group, the ornithischians, as well.
Bakker’s larger purpose was to add further evidence that dinosaurs were warm-blooded. Birds, with their enormous energy and oxygen demands related to flying, were thought to have evolved the air sac system as a way to satisfy the metabolic demands of their endothermy. Thus, if Bakker could show that dinosaurs had this respiratory system too, his arguments about the presence of endothermy in dinosaurs carried more weight. He even makes a comment about “richly oxygen-ated” Mesozoic air, supposing (when he wrote this in 1986) that oxygen levels were higher than present-day levels.
Following Bakker, other dinosaur workers took up the call and the specific case of air sacs being present in sauropods was most recently (and most forcefully) espoused by Berkeley dinosaur paleontologist Matt Wedel in 2003. But by far the most eloquent and substantial arguments for the air sac system being present in the dinosaurian precursors to birds came from Gregory Paul, who in 2002 published his massive and lavishly illustrated Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. In 460 pages, Paul looked at every aspect of the dinosaur to bird transition and the evolution of the air sac system in saurischian dinosaurs, long before the evolution of the first bird, was dealt with in many pages of intricate detail. We need to look at his arguments.
Paul makes many points, several are central to the argument. First and foremost, he points out that the air sac system in flying birds must show adaptations to allow (or at least not impede) flight. He also noted that nonflying birds have air sac systems anatomically distinct from flyers. Thus, the bones of dinosaurs would not necessarily show all the adaptations for air sacs seen in flying birds—that attainment of flight itself would require adaptations to the air sac system not required by nonflyers. Here is the list of characteristics that he finds associated with the air sac system:
Pneumatic bones, especially in the vertebral column.
Shortening of the trunk of the body.
Shortening of the first dorsal ribs.
Elongation and increased mobility of posterior ribs. This mobility is enabled by the presence of ribs with double heads at their ends.
Uncinate processes on several of the ribs (these are small, hook-shaped bones attached to the trunk ribs).
A hinge joint making up the attachment of the ribs with the sternum.
EVOLUTION OF THE AIR SAC SYSTEM
The complex air sac-lung system found in birds had to have evolved from a reptilian, sac-like lung. Here is the pathway envisioned by Paul in his 2002 book.
The first of the so-called archosaurs were the primitive late Permian through early Triassic reptilian group (that we have called diapsids), which would eventually give rise to crocodiles, dinosaurs, and birds. Examples of this group included the quadruped form Proterosuchus (described above as one of the earliest Triassic archosaurs). They would have had a reptilian septate lung. Inspiration may have been aided by a primitive abdominal pump-diaphragm system (more primitive, perhaps, than the system found today in modern crocodiles). Successively, however, evolution of the air sac system may have progressed fairly rapidly. By the time the first true dinosaur was seen in the middle Triassic part of the air sac system may have been in place.
The most primitive theropods from this time (the first dinosaurs) do not show bone pneumatization, but Paul suggests that the lung itself may have become inflexible and smaller—a bird-like trait based on rib anatomy. The ribs also become double headed, showing that the rib cage itself was capable of a great ventilation capacity. Perhaps as a consequence of going bipedal, these dinosaurs may have switched from the more primitive abdominal pump system to the first air sac system—one with only the abdominal air sac found in modern-day birds. Soon after, descendents of these first dinosaurs, forms such as the wellknown, upper Triassic Coelophysis, showed the evidence of bone pneumatization, consistent with the proposal that more air sacs had evolved, this time in the neck region. With the Jurassic forms such as Allosaurus, the air sac system may have been essentially complete (but still much
different from the bird system, modified as it has been for flying, for even the modern-day flightless birds came from flyers in the deep past), with large thoracic and abdominal air sacs. Yet holes in bones do not an air sac system make, if I may paraphrase Yoda.
By the time Archaeopteryx had evolved in the middle part of the Jurassic, there may have been a great diversity of respiratory types among dinosaurs, some with pneumatized bones, some without. There also may have been a great deal of convergent evolution going on. For instance, the extensive pneumatization in the large sauropods studied with such care by Wedel may have arisen somewhat independently from the system found in the bipedal Saurischians.
Paul considers the evidence at hand as proof of a progressively more complex air sac system appearing in middle Triassic to Jurassic dinosaur lineages. He summarized this view in his 2002 book:
One could hardly ask for a better pattern of incremental evolution progressing to the avian skeletal features needed to operate respiratory air sacs. This fact reinforces the case for pre-avian pulmonary air sac ventilation in predatory dinosaurs. No evidence for progressive evolution of a pelvis based diaphragmatic muscle pump (the system found in modern crocodiles) has been presented.
Yet for all these arguments what the paleontologists had were a series of holes in bones, for in no case was a fossil air sac to be found (nor was one expected to be found, of course). And as might be expected, it was not long before a spirited opposition sprang into action. The leader of that opposition was the already introduced John Ruben. During the 1990s he began an extended debate with the advocates of a pre-avian air sac system in general and Greg Paul in particular.
John Ruben and various coauthors came to an opposite conclusion about almost every aspect of what might be called the “air sac in dinosaurs” hypothesis. And they went well beyond even that. In a summary paper published in mid-2005, Ruben and three coauthors proposed that dinosaurs were ectothermic, as were the earliest birds. According to this idea, birds gained warm-bloodedness and the air sac system only with the evolution of flight, and thus warm-blooded, bird-lunged (air sac) birds may not have evolved until the Cretaceous Period, many millions of years after evolution of the first birds. Ruben and his colleagues proposed that dinosaurs possessed simple, septate
lungs that were ventilated with the same system now found in crocodiles—a hepatic air pump diaphragm, that operates by muscles attached to the pelvis. While admitting that some dinosaurs had pneumatized bones, Ruben does not think this is evidence of air sacs. But if not air sacs, what system was used by the dinosaurs? According to Ruben and his colleagues, we have only to look at respiration in crocodiles to see how—and with what organs—dinosaurs breathed.
We have already described the crocodilian system, called the hepatic (liver) piston pump. Like mammals, a diaphragm system inflates the lungs, but, unlike our system, the crocodiles move their entire liver region, as their diaphragm muscles attach partly to the pelvis and partly to the soft tissue of the liver itself along a broad band. The entire liver is pulled back toward the pelvis like a piston, and in so doing the lungs inflate with air. But a piston must fill its cylinder, and such a system would by necessity have a partition right across the body cavity—essentially a subdivision of the forward or thoracic part of the interior of the body from the posterior or abdominal region. Crocodiles have this, and Ruben et al. think that dinosaurs breathed in the same way. They also cite aspects of dinosaur skeletons that seem to preclude the possibility of abdominal air sacs at least. As we have seen, the ribs of birds are capable of extensive movement because of a hinged contact between the posterior ribs and the sternum (the breastbone).
As a further nod to the crocodiles, the Ruben camp does not support warm-bloodedness in dinosaurs. Thus, their views could hardly be more different from the air sac dinosaur endothermy advocates. Voila—there could hardly have been a better recipe for controversy. Unfortunately, neither camp could land any knockout blow to the other’s prime hypothesis based on the evidence at hand. And that is why the discovery of exquisitely preserved feathered dinosaurs from China brought the controversy to a boil.
ENTER THE CHINESE DINOSAURS
The late-twentieth-century discovery of exquisitely preserved bipedal dinosaurs from a spate of quarries in China of early Cretaceous age seemed like exactly the evidence needed to conclude the air sac debate. A requirement of the hepatic piston system, as we saw in the previous
paragraph, is a separation of the body cavity into forward and rear parts by some sort of partition. One of these newly collected specimens, named Sinosauropteryx, showed this type of partitioning—according to Ruben et al. in 1997 and repeated in a review published in Science in 2005. They saw a dark carbonized region in a position within the body cavity where just such a partition is found in crocodiles and because of this they argued that the case was closed—and crowned themselves victors. Paul, of course, had a different view. He looked at this specimen and noted that it had been broken during its collection into many individual pieces and that the required reconstruction of the fossil may have produced the critical black region and thus (in his opinion) could not be cited as evidence of the hepatic piston system. Paul then cited an even more elegantly preserved specimen of a small dinosaur known as Scipionyx, which, according to him, shows the presence of a fossilized air sac!
So back and forth, statement and rebuttal, at this writing the battle rages still (although one side does not seem to know that it has lost). But as noted above, none of the participants considered the effect that potentially low oxygen may have had on the evolution of the early dinosaurs. In fact, Paul, like Bakker before him, cited an older (and now discredited) reference to the Mesozoic having had higher oxygen than now, based on findings of air bubbles trapped in amber. But results of the GEOCARBSULF work indicating oxygen levels during these evolutionary events tell a very different story. It may have been that this prolonged period of low oxygen, more than any other factor, caused what can be called the Triassic Explosion—one part being the evolution of dinosaurs. That said, could the dinosaur air sac question be resolved? Ruben is correct in stating that the presence of pneumatic bones does not “prove” the existence of air sacs in dinosaurs. (Very few scientists ever prove anything.) But the progressive increase in pneumatization in the saurischians strongly supports the theory of air sacs in dinosaurs.
AIR SAC CONTROVERSY RESOLVED?
A really good paper in Nature or Science can tip the scales of any controversy, because publication of any controversial topic in either of
these journals occurs only after rigorous review. A paper in Nature from July 2005, titled “Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs,” by paleobiologists Patrick O’Connor and Leon Claessens brought new morphological data to the controversy and must have felt like a stake in the heart to dear John Ruben.
The paper did not stake out new territory. It looked at existing lines of evidence in a new way, using a nearly unbelievable amount of data collection to support the contention that saurischian dinosaurs (all except the very first, that is) most probably had the avian air sac system. Here is what O’Connor and Claessens did. They looked at birds—a lot of different birds, more than 300, in fact. They looked at birds in more detail than anyone ever has. And by “looking,” the two scientists poked, prodded, filmed, dissected, and injected vast quantities of gooey plastic into the respiratory systems of so many different birds that at the end of the paper they had to plead that all the poor bird “subjects” were dead or dying anyway (of course, they were dead or dying after having their air sacs filled with rubber). But the most amazing thing emerged. The avian air sacs are way more voluminous and complicated than anyone had suspected. By injecting the air sacs with latex, each tiny evagination of an air sac into the bird’s skeleton could be wonderfully observed and characterized. For perhaps the first time, the real relationship of air sac to bone—the pneumatization that we have talked about for so many pages now—could be observed. They even attached a movie of bird respiration, as observed with radiography, in the online version of their paper—thus the birds. But the crux of the paper was in showing that the very specific shapes required for placing parts of the air sacs in bird bones are found in saurischian dinosaur bones. Not just holes in dinosaur bones, but the same shapes of holes in the same (or homologous) bones.
The folks arguing that there was no air sac system in dinosaurs have not denied that dinosaur bones had holes in them. They said the holes were there all right but that they were adaptations simply for lightening the bones. But there comes a point when an argument finally collapses under the weight of too great a coincidence. Like a bad movie in which the plot depends on some coincidental happenstance
far too outrageous for ordinary life, here we would have to accept the coincidence that the same shapes used for lightening bones are also optimal for storing bits of respiratory structure.
The final bit of thunder in O’Connor and Claessen’s paper was in its description of the holes found in an exquisitely preserved early saursichian dinosaur. Because of its importance in the most crucial debate of these middle chapters we can quote from the two authors:
Here we report, on the basis of a comparative analysis of region-specific pneumaticity with extant birds, evidence for cervical and abdominal air-sac systems in non-avian theropods (saurischians), along with thoracic skeletal prerequisites of an avian-style aspiration pump (the air sac respiratory system). The early acquisition of this system among theropods (bipedal saurischians, the first dinosaurs to evolve) is demonstrated by examination of an exceptional new specimen of Majungatholus atopus, (a primitive saurischian dinosaur) documenting these features in a taxon only distantly related to birds. Taken together, these specializations imply the existence of the basic avian pulmonary Bauplan (body plan) in basal neotheropods, indicating that flow-through ventilation of the lung is not restricted to birds but is probably a general theropod characteristic.
A blow had been struck for dinosaurs (or saurischians at least) having the avian, air sac respiratory system. But was it a knockout punch, the kind of data that ends a debate? Or was this the start of another inconclusive round of partisan clinching? The new study made clear that the Ruben group had made a crucial error in asserting that the bone holes in birds only contained the cervical or neck air sacs. For instance, in an influential 2003 paper, Ruben and coauthors Terry Jones and Nicholas Geist wrote:
Pneumatization of the avian skeleton, with the exception of the long bones of the hindlimbs in a small subset of birds, is limited to the axial skeleton and forelimbs and results from invasion by the anterior (cervical and clav-icular) air sacs but is not linked to respiratory function or specific lung morphology. (I have added the italics.)
“Wrong!” argued O’Connor and Claessens. Their exacting observations on living birds showed that holes (pneumatization) in the bones of the middle part of the bird spine were caused by invasion of a different set of air sacs than those suggested by Ruben et al.—most crucially sacs found in the abdominal regions—and which are indeed importantly linked to respiratory function and the very specific lung
morphology. And the bipedal saurischians showed a virtually identical set of holes in the same bones.
What about the ribs in the saurischians? Would they have the mobility necessary to allow inflation of the air sacs? This too seems to be answered by the two authors. They found that the posterior ribs showed a change in attitude that made the air sac system permissible. Also potentially involved are the “gastralia,” free-floating bones found in some dinosaurs.
Perhaps the greatest contribution of this paper was in pointing out that the entire community has misunderstood which air sacs penetrate which bones in birds. Now it is understood that it is the sacral, or tailward, parts of the body that are most important in producing the characteristic “bird-breathing” pattern. So it seems that the case is closed—at least for saurischians. For the ornithischian dinosaurs there is little evidence for the air sac system, and as we shall see, this meshes well with their distribution in time. During the Jurassic times of very low oxygen they were minor elements of the fauna. It was not until the great oxygen rise of the late Jurassic through the Cretaceous that this second great group of dinosaurs became common.
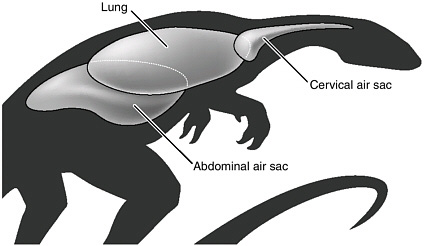
Air sac system in a saurischian dinosaur, as envisioned by O’Connor and Claessens intheir 2005 Nature paper. The cervical (neck) air sacs are present. But also shown are the mid-body and abdominal systems, previously only surmised to be in these kinds of dinosaurs. It appears that the saurischians, like modern birds, were superbly adapted for low oxygen, and this probably accounts for their success.
TRIASSIC- MIDDLE JURASSIC DINOSAURS: A NEW KIND OFANIMAL WITHOUT MODERN COUNTERPART
It is time to come to the heart of the matter about dinosaurs—or at least the earliest forms. The most important questions are whether or not they were warm-blooded and what kind of respiratory system they had in terms of lung morphology, circulatory system morphology (especially the heart), and physiological adaptations for low oxygen. First, let’s look at the environmental context of their first evolution and existence.
The Triassic was a time of low oxygen—which we can find today at high elevations—but of atmospheric gas pressure equal to, or perhaps even exceeding, that of today. This is beyond our experience. It was a time of very high carbon dioxide. Our species has never experienced this (yet we are trying to make it so). It was also a time when the climate of the planet was hot—much hotter than today: no icecaps, no glaciers even in the mountains, and heat from equator to pole. Is it any surprise, then, that the animals of that time may have been unlike anything of the modern-day?
Perhaps the best reference for all things dinosaurian comes from the marvelous, weighty tome The Dinosauria (Second Edition). Edited by David Weischampel, Peter Dodson, and Halszka Omolska, this compendium has data that are currently being mined to show many trends, and some of these data have already been used in the pages above and the pages to come. But in addition to raw data on dinosaur ranges and geographic occurrences, there are summary chapters by leading specialists on all of the issues invoked above, the only difference being that none of the authors takes into account the radically low-oxygen levels during the time of the dinosaurs’ first evolution, conditions that carried on for many tens of millions of years of the Mesozoic. On the twin and central issues of metabolic rate (endothermy versus ectothermy) and respiratory adaptations, there are assured stances that we should look at.
Phylogenetically, dinosaurs are positioned between crocodiles and birds, the former cold-blooded with hepatic piston lungs, and birds with warm blood and the air sac system. It is thus fair to ask where dinosaurs sit in terms of these two end members: Were they closer to
crocodiles or closer to birds based on morphological evidence? And which of the adaptations—cold blood with relatively simple tidal lungs or warm blood with radically transformed and efficient (for low oxygen) lungs did the first dinosaurs possess?
With regard to metabolism, the chapter by dinosaur experts Kevin Padian and Jack Horner points out that dinosaurs were probably more than “damned good reptiles,” citing the rapid growth rates observed in dinosaurs. But were they warm-blooded? Birds are, and it is inferred by Padian and Horner that the direct ancestors of birds, the nonavian bipedal saurischians of the middle Jurassic, may have already evolved this characteristic. But by the time of the first birds (Archaeopteryx) of the late Jurassic, oxygen levels had dramatically risen from their late Triassic nadir. The world was a very different place from the Triassic world, it was one closer to our own, where oxygen was no longer a limiting factor for metabolism.
As far as lungs and respiration go, the chapter by Anusuya Chinsamy and Willem Hillenius took a supposedly dispassionate look at the possibility of an air sac system in dinosaurs that ended up sounding very much as if it had been written by John Ruben. They took the conservative route—no air sacs. And what about the possible presence of air sacs? We know that air sacs ultimately evolved in birds and that the presence of bone pneumaticity makes a strong case for the air sac system. A fully evolved air sac system would have been the best way to deal with the low-oxygen conditions at the time of the dinosaurs’ first appearance and early history, from the late Triassic to the upper Jurassic. But even the true believers, such as Gregory Paul, acknowledge that the basal dinosaurs such as Herrerasaurus showed no bone pneumaticity that would argue for the air sac system and that, if it were present, air sacs in these first dinosaurs would necessarily have been abdominal, thus leaving no fossil record. The evidence for or against air sacs in dinosaurs comes down to rib morphology, rather than bone pneumaticity. As we have seen, the fantastic air sac system in birds can work because their ribs are highly mobile, being jointed or hinged in such a way that would allow the required pattern of avian respiration. Another possible breathing pattern has been proposed, one in which the dinosaurs used dermal ossifications in the abdominal wall to ventilate
an avian-style air sac system. This kind of breathing pattern, called cuirassal breathing, could be used to inflate any air sacs and thus might be evidence that air sacs were present. But again, this system, if it were present, seems less developed in the first bipedal saurischians than in later ones.
Where does this lead us? What systems would be optimal and what systems seem allowable by the osteological evidence of the first dinosaurs? First, metabolism. A warm-blooded dinosaur would have an advantage over a cold-blooded dinosaur—in our world. All modern (cold-blooded) reptiles have to warm up at the start of the day, and thus there is little early-morning activity, other than behavioral movement, in order to acquire heat from the external environment. If the first bipedal dinosaurs—all predators—did not have to do this, they would have been able to forage freely on the slower ectotherms in the cooler morning or nighttime hours. But what is the price for this? At rest, all endo-therms use as much as 15 times the amount of oxygen as do ectotherms (there is a 5 to 15 times range based on experimental observation). In our oxygen-rich world this is not a problem for the warm-blooded animals. So much oxygen is available that there is no penalty. But in the oxygen-poor mid-Triassic, such was surely not the case. And the energy and oxygen necessary for endothermy would not have been necessary if the dinosaurs moved toward large size. With larger body size, the ratio of surface area (from which heat is lost) to body volume becomes increasingly favorable. Truly large-sized animals could have remained essentially homoeothermic in their environment even during cooler nighttime temperatures. While a lizard rapidly loses body heat in a cooler night, a 100-pound reptile does not. And the conditions of the Triassic may have been such that, thanks to highly elevated carbon dioxide levels, greenhouse heating may have kept the temperatures virtually equal day and night—and hot to boot. The Triassic climate was one suited for reptiles—hot. That heat would actually have been a problem for very large endotherms. Large dinosaurs (greater than a ton, such as most sauropods) would have overheated in even moderate temperatures, and the Triassic environment was anything but moderate. So here it is suggested that cold-blooded dinosaurs would have been a condition actually more favorable than warm-bloodedness for dinosaurs, mainly because of the large differ-
ence in oxygen needed while at rest. In the Triassic and into the Jurassic, ectothermy would not have relegated dinosaurs to a sluggish life style. With the absence of nasal turbinals characteristic of modern-day endotherms, the case for ectothermy is stronger than that for endothermy.
And what of lungs? The optimal system would have been the air sac system. Its obvious superiority in today’s low-oxygen environments (at high altitude) would let any animal with such a system have greater oxygen delivery per breath compared to any other kind of lung. But there had to have been a long evolutionary history to arrive at this condition, and since crocodiles, which shared a common ancestor with the first dinosaurs, show no adaptations along this line, the evidence at hand does not support air sacs in the first dinosaurs—rather just the opposite. Neither does the osteological evidence support the hepatic piston respiration favored by John Ruben. The most parsimonious conclusion is that the first dinosaurs and perhaps all dinosaurs, had septate lungs—perhaps highly evolved and efficient septate lungs with subdivisions beyond anything extant today, but septate lungs rather than air sac lungs nevertheless. Further adaptation to low oxygen would have been accomplished by a four-chambered heart with complete separation of venous and arterial blood and perhaps by more red blood cells and other physiological adaptations. Perhaps, too, some of the peculiar bone structure observed by dinosaur bone experts is evidence of larger marrow regions involved in red blood cell formation.
What are we left with? A kind of animal unknown on Earth today. An ectotherm with phenomenally rapid growth rates and a lung system that, while inferior to the best of modern-day birds, was more efficient at extracting the thin oxygen available than those of other denizens of the day. Superiority of the dinosaurs in the latest Triassic and then into Jurassic through the Cretaceous was made possible by being better than everyone else. A return to those times might be surprising indeed, with animals showing behavior that is not mammalian or avian, not a sluggish existence but something in between. Perhaps the earliest dinosaurs were something like lions, sleeping 20 hours a day to conserve energy because of the low oxygen, but when hunting doing so actively, more actively than any of their competitors, which would have included the nondinosaur archosaurs, the cynodonts, and
the first true mammals. All they needed to be was better than the rest. Clearly they were.
The question of metabolic type may also be compromised by the terms used to describe various possibilities. Dinosaur icon Jack Horner certainly thinks so. Metabolic complexes may have been far more diverse than our simple subdivision into “endothermy” and “ectothermy.” While modern birds, reptiles, and mammals are put into one of these two categories, Greg Paul notes that there are many kinds of organisms that can generate heat in their bodies without external heat sources. He includes large flying insects, some fish, large snakes, and large lizards in this camp. Such animals are endotherms but not in the mammalian or avian sense. There may have been many kinds of metabolism in the great variety of dinosaurs that existed.
BACK TO THE SEA
There are other clues than dinosaur bones to the nature of life on Earth and the challenges it faced during the low-oxygen times of the Triassic. Part of the Triassic Explosion was a diversification of reptiles returning to the sea. Many separate lineages did this, and the reasons why this happened may be tied up in the problems posed by the hot low-oxygen Triassic world.
Until now we have stressed respiratory adaptations in various animals. As we have seen, the kind of respiration used by an animal has consequences far beyond simply acquiring oxygen. Oxygen is necessary to run metabolic reactions in animals; it enables the chemical reactions that are life itself. But as in a chemistry experiment, several factors control the reactions themselves. One of the most important is temperature. Metabolic rate is the pace at which energy is used by an organism. It is far higher in endotherms than in ectoderms. But even in the same organism, the metabolic rate is directly and importantly influenced by temperature to a surprising degree. In a 2005 review of animal metabolism, physiologist Albert Bennett of the University of California at Irvine noted that one-third to one-half of all energy expenditure by an animal is used simply for staying alive through activities such as protein turnover, ion pumping, blood circulation—and breathing. Other required activities, such as movement, reproduction,
feeding, and so forth are in addition to this baseline energy expenditure. If metabolic rates go up, so too does the need for oxygen, for the chemical reactions of life are oxygen-dependant. The key finding is that metabolic rates double to triple with each 10-degree temperature rise. The consequences of this in a world that had less oxygen available than now but warmer average temperatures would have been major. Thus, it has been argued here that the large dinosaurs were ectothermic, thus enjoying the best of all worlds (or at least making the best of a very, very bad world, one with no modern counterpart). The enemies were low oxygen and high heat.
There is no direct link between oxygen levels in the atmosphere and temperature. But there is a direct link between temperature and carbon dioxide, the well-known greenhouse effect. And, as we saw in Chapter 2, levels of oxygen and atmospheric carbon dioxide are roughly inverse: when oxygen is high, carbon dioxide is low and vice versa (but not without exception). Thus, there were many times in the past with low oxygen and high carbon dioxide, and thus it was hot. What a double whammy! In a low-oxygen world that is hot, animals lose in both ways—they need more oxygen than in a cool world to run their now faster metabolic reactions but have less oxygen available in the atmosphere!
We have seen many solutions to deal with low oxygen. One is obviously the simple solution of staying cool. Some solutions to staying cool, or cool enough, are physiological; some are behavioral. One of these is morphological, physiological, and behavioral: it is to return to the sea, for even in the hottest world of the past, the ocean would be essentially cooler in terms of physiology. For this reason, perhaps, many Mesozoic land animals traded feet for flippers or fins and returned to the sea at a prodigious rate.
Hypothesis 8.2: In times of higher global temperature but loweratmospheric oxygen, an increasing proportion of tetrapoddiversity is composed of animals that re-evolved a marine lifestyle.
Who has not been struck by the wonders of the Mesozoic, at the marvels posed by those tetrapods that returned to the sea? In the Tri-

Reconstruction of the large Triassic ichthyosaur Cymbospondylus. The ichthyosaurs represented the most extreme body plan change for a once terrestrial group that returned to the sea. The low oxygen on land may have been a major impetus of this.
assic there were giant ichthyosaurs and seagoing tetrapods such as placodonts; in the Jurassic the ichthyosaurs remained and were joined by a host of long- or short-necked plesiosaurs; and in the Cretaceous the ichthyosaurs disappeared, to be replaced by large mosasaurs. The existence of marine tetrapods is no surprise in our whale-enriched world, but the surprise to me was the sense that there were so many kinds back then. This suspicion was finally confirmed with the important research of marine reptile expert Nathalie Bardet, who in 1994 published a review of all known marine reptile families of the Mesozoic.
My reaction back then was a simple “Yes, there were lots of them,” with no further interest at that time. But we returned to this data set when my colleague and research partner Ray Huey suggested to me that the high heat of the early Triassic through Jurassic would have been an evolutionary incentive for some number of reptiles to go back into the sea. We can now test this hypothesis using new data on the number of dinosaurs and the number of marine reptiles (as seen in the previously unpublished graph below). This graph indicated that there is a very interesting and inverse correlation between Mesozoic oxygen levels and the number of marine reptiles. When oxygen was low, the percentage of marine reptiles was high. But as oxygen rose, the proportion of tetrapod families that were fully aquatic markedly dropped. It may not be that the absolute number of marine forms decreased as

Percent of atmospheric oxygen (dotted lines and black squares) plotted against the percentage of marine Mesozoic tetrapod families (black lines and black circles: those vertebrates that returned to the sea, such as ichthyosaurs, plesiosaurs, and mosasaurs). Thisgraph supports the hypothesis suggested here that the number of vertebrates returning tothe sea increased during high-temperature, low-oxygen times. The cooler water wouldhave enhanced survival in a low-oxygen world. This regression analysis of these data ishighly significant (R2=.52). Previously unpublished.
much as it was that the number of terrestrial dinosaurs markedly increased. The figure demonstrating these results is shown above.
FROM LOW-OXYGEN AIR
Let’s now return to the question of why there were dinosaurs. This question can now be answered in multiple ways. There were dinosaurs because there had been a Permian mass extinction, opening the way for new forms. There were dinosaurs because they had a body plan
that was highly successful for Earth during the Triassic. But perhaps these generalizations do not cut to the heart of the matter.
Chicago paleontologist Paul Sereno, who has unearthed some of the oldest dinosaurs and has made their ascendancy a major part of his study, looks at the appearance of dinosaurs in another way. In his 1999 review The Evolution of Dinosaurs, he noted: “The ascendancy of dinosaur on land near the close of the Triassic now appears to have been as accidental and opportunistic as their demise and replacement by therian mammals at the end of the Cretaceous.” Sereno suggested that the evolutionary radiation following the evolution of the first dinosaurs was slow and took place at very low diversity.
This is quite unlike the usual pattern seen in evolution when a new and obviously successful kind of body plan first appears. Usually there is some kind of explosive appearance of many new species utilizing the new morphology of evolutionary invention in a short period of time. Not so with the dinosaurs. Sereno further noted that the dinosaurian radiation, launched by 1-meter long bipeds, was slower in tempo and more restricted in adaptive scope than that of therian mammals.
For millions of years, then, dinosaurs and other land vertebrates remained at relatively low-standing diversity, a finding that Sereno and others continue to find perplexing. In my view, this question can now be answered. Earlier we showed that there appears to be a correlation between atmospheric oxygen and animal diversity: times of low oxygen saw, on average, lower diversity than times with higher oxygen. It appears that the same relationship held for dinosaurs. To formalize this:
Hypothesis 8.3: Dinosaur diversity was strongly dependent onatmospheric oxygen levels, and the long period of low dinosaurdiversity after their first appearance in the Triassic was dueto the extremely low atmospheric oxygen content of the lateTriassic.
Support for this hypothesis comes from our previous analysis of animal diversity in times of low oxygen versus high oxygen. Low-oxygen times apparently stymied the formation of many individuals (while at the same time stimulating experimentation with new body plans to deal with the bad times). This relationship has been demon-
strated for marine animals, and it seems to hold for dinosaurs and other vertebrates as well. Let’s examine further evidence in support of this hypothesis in the next section.
ON THE NUMBER OF DINOSAURS
Support for this hypothesis can be found by comparing two new data sources: the latest GEOCARBSULF results for the Triassic through Cretaceous and a new compilation of dinosaur diversity through the same sampling period.

Atmospheric oxygen percentage plotted against number of dinosaur genera. This figuresupports the hypothesis that higher oxygen supported a higher diversity of dinosaurs.Part of the reason for this may be due to the fact that rising oxygen levels opened upmore habitable areas at altitude, a prediction from Huey and Ward, 2005. Previouslyunpublished.
Recent work by David Favkosky and colleagues, published in 2005, has provided our best estimate yet of the number of dinosaurs during the three periods of the Mesozoic. They show dinosaur genera staying roughly constant from the time of the first dinosaurs in the late middle and late Triassic through most of the Jurassic. While surely mitigated by collecting biases, the sheer number of identified dinosaur skeletons probably ensures that their overall trend is real.
It is not until the latter part of the late Jurassic that dinosaur numbers started to rise significantly, and this trend continued inexorably to the end of the Cretaceous, with the only (and slight) pause in this rise coming in the early part of the late Cretaceous. This slight drop may be due to the very small number of known localities of this age yielding dinosaurs. By the end of the Cretaceous (in the Campanian Stage) there were many more dinosaurs than during the Triassic to upper Jurassic. What was the cause of this great increase?
The figure above certainly suggests that changing oxygen levels were coincidental with changing dinosaur diversity. This is probably more than coincidence. Through the late Triassic and first half of Jurassic, dinosaur numbers were both stable and low. While originating in the latter part of the Triassic, they stayed relatively few in number until a moderate rise at the end of the period—a rise that seemed to coincide with the end-Triassic mass extinction itself. Gradually, if the oxygen results from GEOCARBSULF are even approximately correct, oxygen rose in the Jurassic, hitting 15 percent or more in the latter part of the period. It was then that the number of dinosaurs really began to increase. It was also at this time that the sizes of dinosaurs increased, culminating in the largest dinosaurs that ever evolved appearing from the latest Jurassic through the Cretaceous. Oxygen levels steadily climbed through the Cretaceous and so too did dinosaur numbers, with a great rise found in the late Cretaceous, the true dinosaur heyday. There were surely many other reasons for this Cretaceous rise. For instance, in mid-Cretaceous times the appearance of angiosperms caused a floral revolution, and by the end of the Cretaceous the flowering plants had largely displaced the conifers that had been the Jurassic dominants. The rise of angiosperms created more plants and sparked insect diversification. More resources were available in all ecosystems, and this may have been a trigger for diversity as well.
TRIASSIC-JURASSIC MASS EXTINCTION
Let’s finish this chapter with one last bit of evidence pertaining to the kind of respiration present in late Triassic animals. One of the striking new findings of the latest GEOCARBSULF has been the level of Triassic oxygen. Only several years ago the minimum oxygen levels of the past 300 million years were universally pegged at the Permian-Triassic boundary of 250 million years ago. But that time of low oxygen has been substantially moved and now may correspond more closely than previously thought with the Triassic-Jurassic boundary of 200 million years ago. Thus, we are confronted with the possibility that oxygen was lower in the late Triassic than in the early part of the period—perhaps as low as 13 percent of the atmosphere at sea level, or much less than modern-day levels. This time corresponds to one of the major changes of the Triassic: the winnowing out of most land vertebrates, with the exception of the first dinosaurs, the saurischians—creatures with pneumatized bones. This realization comes from new data compiled for this book by one of my grad students, Ken Williford, who very painstakingly went through the literature describing the various Triassic vertebrates and their ranges. The results of this long search are shown in the diagram below.
In the figure below, the Triassic-Jurassic mass extinction, one of the five most deadly mass extinctions of the past 500 million years, is shown as the horizontal black line in the middle of the figure. Here the saurischian (dinosaurs) are considered to have had air sacs.
The data gathered for this figure showed that every group except the saurischian dinosaurs was undergoing reduction (or at best, maintaining roughly equal diversity) in the time intervals leading up to and after the Triassic-Jurassic mass extinction. The groups with the simplest lungs (amphibians and thecodont reptiles) fared the worst and many groups that had been very successful early in the Triassic, such as the phytosaurs, underwent complete extinction. Both amphibians and thecodonts probably had very simple lungs inflated by rib musculature only. Mammals and advanced therapsids of this time, probably both having diaphragm-inflated lungs, did better, but crocodiles, presumably with abdominal pumps, did poorly. The success of the saurischians may have been due to a multitude of factors (food acquisition, tem-
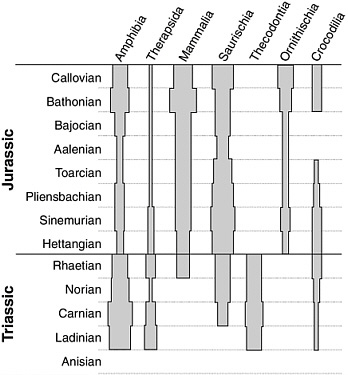
Diversity of tetrapod genera from the middle part of the Triassic to the middle part ofthe Jurassic, a time interval of some 60 million years, with hypothesized breathingsystems for tetrapod orders. Note that of all the orders shown, only the saurischiandinosaurs increase in diversity leading up to and after the Triassic-Jurassic mass extinction. Their then unique air sac system may have been the prime reason for this.Previously unpublished.
perature tolerance, avoidance of predators, reproductive success), but my conclusion is that this group was unique in possessing a highly septate lung with air sacs that was more efficient than the lungs of any other lineage and that in the very low-oxygen world that occurred both before and after the Triassic-Jurassic mass extinction, this respiratory system conveyed great competitive advantages. Under this scenario, the saurischian dinosaurs took over the planet at the end of the Triassic and kept that dominance well into the Jurassic because of superior activity levels, which was related to superior oxygen acquisitions.
We now know that, alone among the many kinds of reptilian body plans of the middle to late Triassic, the saurischian dinosaurs diversified in the face of either static, or more commonly reducing, numbers
in the other groups. We also know that oxygen reached its lowest levels of the past 500 million years in the late Triassic. Something about saurischians enhanced their survival in a low-oxygen world. In another conversation with John Ruben, I was told that this only shows that the abdominal pump allowed this enhanced survival. But what about the bone pneumaticity shown only by the group that prospered through a very bad time in Earth’s history (at least if you were an air breather). And why did the other groups supposed to have had abdominal pumps do so badly—such as the phytosaurs, which went completely extinct?
The ground truth suggests that a long and slow drop in oxygen culminated in the Triassic mass extinction but that this extinction was really a double event, separated by between 3 and 7 million years. There are few places on land where this time interval with abundant vertebrate fossils can be found. We really do not know the pattern of vertebrate extinction as well as we do for the extinction in the sea. We do not know how rapidly the prominent vertebrate victims of the mass extinction such as the phytosaurs, aetosaurs, primitive thecodonts, tritylodont therapsids, and other large animals disappeared. But by the time that the gaudy Jurassic ammonites appeared in the seas in abundance, leaving behind an exuberant record of renewal in early Jurassic rocks, the dinosaurs had won the world.
What kind of lungs did they have? Here it can be proposed that they had lungs and a respiratory system that could deal with the greatest oxygen crisis the world was to know in the time of animals on Earth. Let’s formalize this:
Hypothesis 8.4: Saurischian dinosaurs had a lower extinctionrate than any other terrestrial vertebrate group because ofa competitively superior respiration system—the first air sacsystem.
Support for this hypothesis comes from the new anatomical findings of an air sac system in saurischians and the newly presented data (above) that saurischians were actually expanding in number across this mass extinction boundary. They emerged—perhaps gasping, but still standing, over the corpses of the simpler lunged—in a world we will look at in the next chapter.