Memory: The Key to Consciousness (2005)
Chapter: 2 Memories of the Here and Now
2
Memories of the Here and Now
We begin our exploration of the human memory system by considering primary memory, a term used by the psychologist-philosopher William James in 1890 to refer to the contents of our immediate, ongoing awareness. Since James’s time, it has been variously called immediate memory, short-term memory, and working memory. The modern version of James’s concept of primary memory originated in research by the British psychologist Donald Broadbent and the American psychologist George Miller, who analyzed the problems of attending to and processing events that occurred simultaneously or in quick succession. Intense experimental study of these problems has produced detailed understanding of consciousness, attention, perception, and memory. More recently, work has started on understanding of the brain systems responsible for these basic human capacities.
The Two-Store Model of Memory
A decade of memory research led to the diagram shown in Figure 2-1. Based on the work of Richard Atkinson and Richard Shiffrin, it is a schematic of how information flows through the various aspects of our declarative memory system, and it is usually referred to as the two-store model of memory (even though it actually consists of three memory stores).
Sensory Memories
Imagine that you are a subject in a psychology experiment in which you are shown an array of letters such as the following:
H T D R
B F Q G
L N S K
You see the array for only 50 milliseconds (one-twentieth of a second), and you have to recall as much of the display as you can.
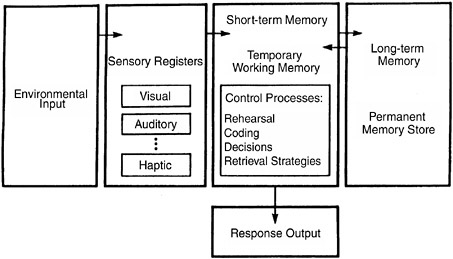
FIGURE 2-1Atkinson and Shiffrin’s model of the short-term and long-term memory systems.
Some people appear to possess a rare ability to take in and retain large amounts of visual information. You, however, are much more likely to be able to recall only four or five letters and their locations. Interestingly, you would perform at about the same level if the experimenter showed you an array of common objects rather than random letters of the alphabet. This apparent limit on perception and memory was originally known as the span of apprehension (“apprehension” in the sense of grasping something). Actually, all of us possess a high-capacity, high-resolution visual memory, one that holds a lot more information than the span of apprehension seems to indicate. This is the good news. The bad news is that for most of us, our visual short-term memory lasts only about a fifth of a second.
How this was first determined is an interesting detective story. In 1960 George Sperling, now at the University of California at Irvine, took a closer look at the span of apprehension and the belief that it is only four or five stimulus objects. He succeeded in showing that immediate visual sensory memory has a capacity much greater than this. Sperling argued that the image of the array might be fading during the few seconds it took the person to report what they had seen (our first example of how the act of remembering can actually cause forgetting). He tested this hypothesis in a simple and elegant series of experiments in which he used a device that projected the array of letters very briefly and then presented a cue to the subject to report only one row of the array (for example, a high-pitched tone to signal recall of the first row, a medium-pitched tone for the second row, and a low-pitched tone for the third row). The cue occurred at various time delays after the image had been presented, ranging from 0.02 second to 1 second. He repeated this over and over with the cue in different random and unpredictable locations in different arrays. The key finding was that if the cue was presented less than 0.1 second after the array had vanished, people could typically report most of the letters in any given row but could not do so if the cue was delayed for 0 .5 second or more. Sperling reasoned that this must mean the memory image of the array contained information about
all of the letters and that the capacity of this form of memory was therefore actually much greater than four or five elements.
Sperling’s experiments are also important because they provide an example of what appears to be a simple decay of information from memory. Visual information enters a sensory register or “iconic” memory where it is held in detail for a brief period. Some of this information is transferred to a short-term memory store if it is attended to, and some may even get transferred to a more permanent long-term memory; but much information from sensory memory is not attended to, is not stored, and is simply lost. (The process of attention and its importance in memory creation are discussed in more detail later in this chapter.)
Visual sensory memory can also be easily interfered with and essentially erased. Suppose you see a row of digits such as 5 3 8 4 flashed on a computer monitor for 30 milliseconds, followed by a blank screen. You will be able to remember all four digits in order almost all of the time. But suppose the row of digits is followed immediately not by a blank screen, but by a row of symbols such as # # # # that occupies the same location on the screen as the digits. Under these conditions, you won’t be able to recite the digits easily. In fact, you might find yourself telling the experimenter that you didn’t even see any digits! This is a simple demonstration of what’s called backward masking, an effect that seems to occur because the symbols overwrite the digit information in the iconic memory store before it can be transferred to the next stage of processing. If you think about it, this seems to say that in ordinary, moment-to-moment vision, information is continuously being erased from iconic memory as new information enters the visual system.
As Figure 2-1 suggests, there are short-term sensory memory stores for other senses as well: hearing (the acoustic store), touch (the haptic store), and smell (the olfactory sensory store). They all seem to have the same general function and properties as Sperling’s iconic memory: They preserve sensory information for very brief periods but are highly susceptible to loss of information from decay and interference.
Are there people with “photographic memories” who can perceive and remember a 3 by 4 array of random letters after a single 50 millisecond exposure? There have been a few reports of such individuals, but they have not withstood scientific scrutiny very well. At the same time, there definitely are individual differences in the capacity of visual short-term memory, and recent research has begun to identify the brain regions where these differences may reside. Working independently, Jay Todd and Rene Marois at Vanderbilt University and Edward Vogel and Maro Machizawa at the University of Oregon tested undergraduate subjects whose average performance ranged from an average of one or two correct to five or more correct in tests of memory for briefly presented arrays of colored dots and squares. Both research teams found that low- and high-capacity subjects differed in levels of brain activity in the parietal cortex, a region of the brain known to be involved in the processing of visual and spatial information.
The Short-Term Store
The concept of memory span was known to psychologists long before the two-store model of memory was developed. It refers to the longest randomly ordered sequence of stimuli—digits, letters, words—that a person can recall in order after one brief presentation. Test someone with the digits sequences shown here, reading them at a rate of two per second. A list
2, 7, 5, 1
2, 7, 5, 1, 4, 8, 9
2, 7, 5, 1, 4, 8, 9, 3, 6, 1, 7
of four digits is easy recalled; a list of seven digits is noticeably harder; and a list of 11 digits is next to impossible to recall perfectly. Young adults have a memory span of seven or eight for randomly ordered digits.
This basic limit on our ability to recall very recent events is represented in Figure 2-1 by the short-term memory or working
memory store, which is thought of as a temporary storage system that has a relatively small capacity. The concept of a short-term memory store was also inspired by the observation that the last few events of a sequence tend to be relatively easy to recall. This is called the recency effect. If you present subjects with lists of 15 common words at a rate of one word per second and have them recall the words as soon as the list ends, in any order they can recall them, then you will get results like those shown in Figure 2-2. Recall of the first few words in the list is relatively good but drops off toward the middle of the list; then recall rises steeply toward the end of the list. This whole pattern is called the serial position effect in immediate recall. According to the two-store model, the recency effect occurs because the last five or six words in the list are still available in the short-term store at the time of the test.
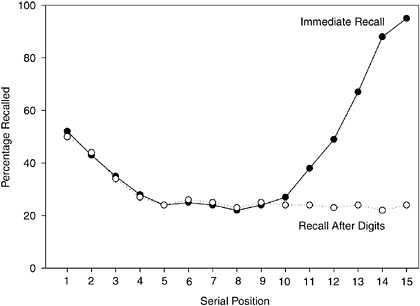
FIGURE 2-2The serial position effect in recall of a sequence of 15 words. The prominent recency effect can be eliminated by a brief distraction such as reciting a list of digits.
An interesting fact about the recency effect in this kind of experiment is that it is very fragile. Suppose we change the experiment so that the subject has to recite just three digits that appear after the fifteenth word and then recall the words. As shown in Figure 2-2, the recency effect disappears almost completely! In this regard, the short-term store is just like the sensory store because its contents can be readily displaced by new incoming information that requires attention and competes for “space” in short-term memory.
This fact has an important practical implication: Being able to recall the contents of short-term store after a short, distraction-free interval does not mean that you will also be able to recall them as easily after a longer interval filled with other events, as many witnesses to crimes or traffic accidents have discovered.
Long-Term Store
We’ll complete our exploration of the two-store model by looking at some of the most important properties of long-term storage or long-term memory. According to the model shown in Figure 2-1, information moves from short-term store to a larger and more permanent storage system. Whether or not this happens depends largely on the process of encoding that is applied to information while it is still in short-term store. For example, suppose in trying to remember a telephone number you’ve just looked up, you repeat it to yourself (an act called “maintenance rehearsal”). This typically succeeds in keeping the information active and accessible in the short-term store, but is a poor strategy for creating a new memory record that will last over time and in the face of distractions. Something more is needed.
George Miller, a pioneer in the study of cognitive psychology and memory, suggested that this “something more” often consists of the process of chunking. Miller described the capacity of short-term memory as “the magical number 7, plus or minus 2.” By this he meant we can hold about seven chunks of information in short-term memory. The word “chunk” here means something like “psychological unit” or “perceptual unit.” Look back at the
example of the letter array used in Sperling’s experiments. It represents 12 chunks of information because the letters, being randomly chosen and arranged, each represent one psychological or perceptual unit. But suppose you were tested in the Sperling experiment with arrays like this:
H O L D
B A R K
L I N E
The number of letters you could recall would now be much greater because, while there are still 12 letters, they represent a much smaller number of chunks (just three familiar words). The same principle applies to memory span. The letter sequence O U T S T A N D I N G consists of 11 letters but only one chunk, and is easily accommodated by short-term memory and easily transferred into long-term memory. This also explains why memory span for meaningful sentences is far greater that for randomly ordered digits or words. You would not have much trouble remembering a 14-word sentence such as the wicked old witch led the two trusting children into the deep dark forest even when it is shown to you word by word at the very fast rate of 10 words per second.
Chunking also seems to be the basis of some kinds of learning. For example, if you are shown a list of 30 unrelated words, one at a time for one second each, you may be able to recall only about six to eight of them if you’re tested right away. But if you are shown the same words a second time, the amount you can recall increases, and will continue to increase every time you are given an additional learning trial until you get to the point where you can easily recall all 30 words. One important reason you’ve been able to overcome the “7 plus or minus 2” limit is that in the course of learning you have formed information-rich chunks, each of which may contain several words. See Box 2-1 for an example of how the chunking principle can be applied to create a memory span of over 70, and for a discussion of memory improvement in general.
BOX 2-1 Can we learn to improve our short-term memory? The answer is yes, with some qualifications. In one experiment the subject was a young man who was an enthusiastic amateur runner. At the beginning of the project he had a normal memory span for digits, about seven. He was given massive amounts of practice in which he applied a special encoding or chunking procedure to the digits sequences. He would try to recode a set of digits into something that was familiar and meaningful: times in foot races of different distances. For example, 3 5 4 5 1 might be encoded as “near world record for a one-mile race,” and this seemed to reduce the five digits to one chunk. With a great deal of practice and effort, he was also able to “chunk chunks” until he got to the point where he could recall up to 70 digits after hearing the sequence just once. What do you suppose happened when he was then given some tests with randomly ordered letters of the alphabet? His memory span was right back to where it had started–around seven letters! This is a good illustration of what strategies for memory improvement most often consist of. Rather than increasing the actual capacity of a memory store, they work by improving the way a person encodes information into long-term memory. This basically means using what you already know—what’s already in your long-term memory—to assimilate new information. Many kinds of mnemonic devices are based on this principle, such as the sentence codes you learned as a child to represent the notes of the treble clef or the rhymes that represent spelling rules. And if it’s ever important for you to learn and never forget which kind of camel has two humps and which has one hump, just remember what you see when the B in Bactrian and the D in Dromedary are rotated 90° to the left. In general, how well new information is stored in long-term memory depends very much on depth of processing, an important concept developed by Fergus Craik and Robert Lockhart of the University of Toronto. A semantic level of processing, which is directed at the meaning aspect of events, produces substantially better memory for events than a structural or surface level of processing. For example, if you respond to each word in a list that an experimenter wants you to remember by thinking about the pleasantness of each word, your memory for the word list will be much better than if you had instead paid attention to the number of letters in each word or to some aspect of their perceptual appearance. Interestingly, the same result occurs if you want to remember faces. As you look at a face, judge the person’s trustworthiness or temperament. Your later recognition of the face will be considerably better than it would be if you had instead focused on facial features only. |
Forming Long-Term Memories
Even with minimal exposure to an event, some information about that event seems to register in long-term memory. An example of this is called the Hebb-Melton effect, in which subjects are given a sequence of memory span tests for random nine-digit sequences such as the following:
4 8 5 2 6 3 7 1 9
5 7 2 9 3 6 2 8 1
8 1 5 7 3 4 2 6 9
4 8 5 2 6 3 7 1 9
The subjects are not told that a given digit list may be repeated (for example, the first and fourth sequences in this example). What is found is that recall of the second occurrence of a given list is better than recall of the first occurrence. In simple terms, some learning or transfer of information into a more permanent form must have occurred when the repeated sequence was first presented and tested, even though the memory task involved would require only transitory short-term memory.
It sometimes seems that all that is required to produce a durable long-term memory is perception of a meaningful stimulus event. This was demonstrated in dramatic experiments by Ralph Haber at the University of Rochester and Lionel Standing at Bishop’s University. In these experiments, undergraduate students were shown a large number of slides of color magazine photos that depicted a wide variety of scenes, people, and objects. Each slide was shown for four or five seconds. A few days later subjects were shown a series of pairs of pictures in which one picture was “old” (seen previously in the experiment) and one was “new” (not previously seen) and they were asked to pick out the old pictures. After viewing 1,000 pictures, they scored an amazing 90 percent correct in this recognition test. They did pretty well even when the “new” test pictures were mirror-image versions of the original pictures!
The memory system of the human brain seems to have an astonishing capacity for this kind of material. In one of Standing’s experiments, subjects were shown 10,000 slides over a two-day period, with each slide exposed for five seconds. The performance of subjects on the recognition memory test indicated that they had retained information for at least 6,600 of the pictures. This doesn’t mean that the subjects were storing highly detailed representations of each picture in memory, but it does suggest that some information of the visual scene in each slide had been stored directly in visual long-term memory.
There is an interesting connection between what is known about the capacity of visual long-term memory and the use of mental imagery in learning and memorizing. Many schemes for rapidly assimilating and retaining information are based on the use of visual mental imagery (see Box 2-2).
A third example of how readily long-term memory can be formed from brief experiences can be found in studies of what are called priming effects. In one experiment, Carolyn Cave had subjects name pictures of common objects as fast as they could. One year later the subjects were retested with some of the pictures they had seen and named a year before, along with some new pictures. She found that subjects were faster at naming the previously seen and named pictures than the new pictures—even when they couldn’t remember having seen the pictures originally. This is astonishing: A brief experience in a memory experiment created memory traces that lasted at least a year.
Attention, Consciousness, and Memory
Psychologists think of attention in terms of how we allocate our perceptual and mental resources to some aspects of a complex environment. If you are reading a book and someone speaks to you, your behavior can range from not hearing the person at all and being aware of (and remembering) what you are reading to being fully aware (and remembering) what was said to you and not remembering what you are reading. Two key assumptions of
BOX 2-2 Some of the most effective methods for memorizing material quickly and effectively are based on use of “the mind’s eye”—our ability to visualize objects and places and to form mental images from them. Some of these methods (mnemonic devices) have been known since antiquity. One is the method of loci (locations). Suppose you want to memorize a list of things you must do or a set of words in a fixed order. The best way to proceed is to form a mental image of each of the things to be remembered as occupying a place along a travel route you already know well. Then, when you have to retrieve the information, just take a stroll (a mental one) along the route. You will find that the things you have to remember come to mind quite easily. Another simple mnemonic system is the rhyming peg word method. Suppose you have to memorize a list of 10 words (cash, doll, band, farm, hill, gang, knee, lime, mail, nose) in a way that preserves information about the order of the words as well as the words themselves. This is not easy to do if you use a learning strategy such as repeating the word list over and over, but it’s a breeze if you use the rhyming peg word system. All you need to do is first learn the “peg words” for the numbers 1 through 10:
The next step is to form a mental image of the words you have to memorize, associating each word with successive peg words. For “one” imagine a gun lying on a pile of cash; for “two” think of a shoe with a doll stuffed in it; and so forth. This system is amazingly effective. It is also easy to learn to use, unlike some mnemonic systems that require a great deal of initial memorization and practice. The use of mental images in mnemonic devices is just one example of the important functions of visual mental imagery in human memory and cognition. The modern study of mental imagery was started in the 1960s by Allan Paivio of the University of Western Ontario in a wide-ranging program of research that led to Dual Code Theory, which argues convincingly for the existence of separate verbal and visual systems for representing information in memory. |
all theories of attention are that there are limits to our attentional resources as well as limits on how well we can divide attention. Think of a flashlight beam that can illuminate only so much space at any one time.
The scientific study of selective attention is another intriguing detective story. How can you determine if someone is attending to something? Typically you see if they remember what they were attending to or supposed to be attending to. Suppose you played two different stories through two earphones of a headset. If you ask people to pay attention to the right ear, they will remember that story but be unaware of the story in the left ear, and vice versa. This led to the notion of a selective filter. The unattended message was “filtered out” before it reached awareness.
This filter model was appealing. But Anne Treisman, a leading authority on human cognition, added a twist. At a certain point in the unattended message spoken to the left ear, the listener’s own name was spoken. The person immediately became aware of the unattended message. Somehow, the unattended message was being evaluated someplace in the brain and was determined to be important or unimportant. This has been called the “cocktail party effect.” You are talking to someone at a crowded and noisy party. You are unaware of the contents of the conversation going on behind you until your name is spoken. Suddenly you attend to that conversation and ignore the person speaking directly to you.
Notice the implication here that the sensory and perceptual systems of the brain can detect, analyze, and filter out information about events in the external environment before you become consciously aware of them. Exactly how this preconscious processing happens is currently a major issue in cognitive neuroscience and brain science, but it is clear from the results of many different kinds of experiments that the processing of sensory information by the brain does not always lead to conscious awareness of an event; it depends, among other things, on how your attentional resources have been deployed.
In one such experiment, subjects saw a rapidly presented
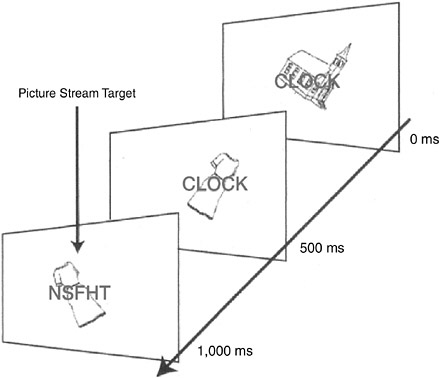
FIGURE 2-3A rapidly presented stream of pictures, words, and random letter strings.
stream of either meaningless strings of letters or familiar words superimposed on pictures (see Figure 2-3) while undergoing a brain scan by MRI. Their task was to detect repetitions of pictures and ignore the verbal material, or vice versa. Normally, the brain activations produced by a random string of letters and by a real word are usually quite different. However, when subjects in this experiment were instructed to attend and respond to the pictures only, the brain activations for the superimposed nonsense words and real words were about the same, even though the subjects were looking directly at the words. (Try looking at one of the pictures without perceiving the word; it seems impossible.) At the end of the experimental session, subjects could remember many of the words if they had been directed to pay attention to them, but had
no memory at all for the unattended words. It seems that focusing of attention on one kind of event (the pictures) precluded processing of the other kind of event (the words)—even though the perceptual and memory systems of the brain would be able under normal conditions to detect and identify the words and to bring them into conscious awareness in a few fractions of a second.
Another striking demonstration of this “inattentional blindness” was conducted by Daniel Simons and Christopher Chabris at Harvard University. Using an experimental procedure first devised by Ulrich Neisser, they had undergraduate subjects look at a video depicting a group of students moving about and passing a basketball around. The subjects were instructed either to keep track of the number of times the ball was passed (the easy task) or to keep separate tallies of the number of air passes and bounce passes (the difficult task). They were not told that something unusual would happen in the middle of the video: A person wearing a gorilla suit entered the scene, paused in the middle of the group of basketball passers, beat on its chest, and then exited (Figure 2-4). What Simons and Chabris found was that at least 35 percent of the subjects did not report seeing the gorilla in the easy task, and more than 50 percent failed to see the gorilla when the counting task was difficult! It is important to understand that in this experiment the person in the gorilla costume was easily visible for several seconds and would always be seen by someone asked to simply look at the video but not required to do any counting. Simons also found that subjects counting basketball passes by students in black shirts were more likely to report seeing the gorilla than subjects who had to count passes by students in white shirts. This is what you would expect if the student observers had “set their attentional filters” to select one kind of stimulus event and filter out others.
Consciousness
Most discussions of attention lead to the issue of consciousness and awareness.

FIGURE 2-4You can fail to see the person in the gorilla costume if your attention is focused on the students passing the basketball around.
Some theorists believe that the primary brain system responsible for consciousness is the cerebral cortex. A compelling case for this view was made by the neuroscientist Larry Weiskrantz at Oxford University in elegant studies of a person with “blindsight.” This individual had extensive damage to the primary visual areas in the cerebral cortex. As far as the patient was concerned, he was totally blind in a large part of his visual field. When presented with a spot of light somewhere in his visual field and asked to point to it, he would say, “what spot of light? I don’t see anything.” If you persist and tell the patient just to point anywhere he will humor you and point at random—except that he doesn’t point at random; he always points accurately to the spot of light. But he doesn’t see it at all; he is not aware of the spot of light and has no working memory of it. But other visual systems in his brain are able to direct his pointing. Such patients also avoid objects. In walking through a room they walk around chairs and other objects all while insisting that they can’t see any of the objects.
As mentioned in Chapter 1, some perception and memory for-
mation can occur without consciousness, at least judging from experiments performed on patients under deep surgical anesthesia. If a list of words is read to them while they are anesthetized, they will later, when awake, have no episodic memory at all of the words or, for that matter, any aspect of the surgery. But if you give them an implicit test for the words, they will show evidence of having “registered” more of the words than control subjects who were not read the words. This priming memory seems to involve auditory-visual association areas of the cerebral cortex. Perhaps awareness or consciousness is what we can describe verbally at a given moment in time—the contents of our working memory.
Where does this leave animals? Evolution works in very small steps to change the structures and functions of animals. If humans have awareness or consciousness, it seems reasonable to suppose that nonhuman animals have a similar capacity. It must have evolved gradually from small beginnings in simpler animals because it has adaptive value. But how can it be assessed in animals, who cannot speak? We can teach them how to tell us. In ingenious experiments, monkeys with damage to the visual cortex showed blindsight. They can be trained to tell us that they cannot see a light, yet they can point to it. These monkeys, like humans, have lost awareness of the visual stimulus.
There is other good evidence that some species have a form of conscious episodic memory much like that of humans. Anthony Wright has studied “list memory” in monkeys using procedures not so different from those used with human subjects. This research has produced evidence for common processes in nonverbal monkeys and verbal humans. For example, monkeys show recency effects and serial position effects (see Figure 2-2) that resemble those of humans in important ways.
When a questionnaire was sent to many neuroscientists asking them to rank animals in terms of degree of consciousness, the results were just what might have been expected. Primates and possibly sea animals ranked highest, then carnivores, then rodents, and so on. The neuroscientists expressed serious doubt
about the consciousness of flies and worms. The result of the questionnaire is of course merely opinion, but here opinion corresponds rather closely with the evolution of the forebrain and cerebral cortex.
Working Memory and Its Brain Systems
As is true for most really important ideas in science, the two-store model of memory has been criticized in virtually every conceivable way, but basics of the model of it have stood the test of time. When Atkinson and Shiffrin first proposed the two-store model of memory some 30 years ago, they described short-term store as a temporary “working memory” that could be used in flexible ways to adjust to the demands of processing information. Over time this idea has become a central one in studies of memory, mind, and brain. For many contemporary theorists, working memory is much more than a small, passive memory store; instead, it is an essential part of conscious awareness, with important connections to attention and mental ability.
Alan Baddeley of Cambridge, England, a leading authority on working memory, characterizes it as the temporary storage of information in connection with performing other, more complex tasks. He proposes a multicomponent system, comprising an attentional system, the central executive, aided by “slave” systems responsible for temporary storage of either visual or verbal material. He coined the very apt terms visuospatial sketchpad for visual temporary storage and phonological loop for temporary storage of verbal speech material. As we shall see, his view appears to correlate well with findings on the brain mechanism of working memory.
The involvement of the prefrontal cortex in working memory in monkeys was first reported by C. F. Jacobsen at an international congress in 1935. Frontal-lobe lesions in the brains of monkeys produced a marked taming effect on the animals. A neurosurgeon, Egon Muniz, was in the audience. He immediately saw the relevance of this effect for humans, and at his clinic in
Portugal he began removing the frontal lobes in humans to treat psychiatric problems. Unfortunately, Jacobsen had also described a severe deficit in working memory following the lesion in monkeys. Thousands of human operations later it was realized that this damaging procedure was of little help in the treatment of mental illness. A most unfortunate chapter in the history of psychiatry, the procedure is no longer used today.
A standard task now used to test short-term memory in monkeys (and human infants) is the delayed response. A monkey is shown two food wells; one is baited with a preferred food, and both are covered with identical objects. An opaque screen is placed between the animal and the objects for a short period of time. The screen is lifted and the animal must reach out and displace the object over the food to obtain it (see Figure 2-5). Note that this is not a visual discrimination as such but rather memory of a location, a kind of spatial short-term memory. Normal animals can learn to perform correctly with delays of many seconds. Destruc-
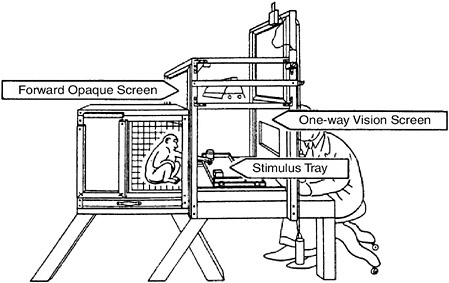
FIGURE 2-5Testing a monkey’s short-term memory.
tion of a localized region of frontal cortex severely disrupts the animal’s ability to perform this task, even at relatively short delays.
In a visual discrimination task, the animal must learn to choose one of two different objects to obtain a reward, that is, to learn to recognize particular objects. Again, normal animals can learn to do this well even with delays of many seconds. Lesions of a different region of prefrontal cortex impair this task, even with relatively short delays.
Much of our current information about the monkey prefrontal cortex has come from two scientists, the late Patricia Goldman-Rakic at Yale University and Joachim Fuster at the University of California at Los Angeles. In addition to lesion studies, they have analyzed the activity of single neurons in a monkey’s prefrontal cortex while the animal is performing delay tasks. They found neurons that increase their firing rates during delay periods, as though they are holding the needed information in working memory.
Important studies by Brenda Milner at McGill University showed that patients with prefrontal damage are greatly impaired in remembering the temporal sequence of events, although memories for the events themselves are not impaired. Thus, Milner would show a patient a series of pictures of paintings, one at a time, and at some point show the patient two paintings and ask which was seen first. Patients with prefrontal damage are much impaired on this task (which is typically easy for nonimpaired people), although they remembered perfectly well having seen the paintings earlier.
If any one term summarizes these rather diverse aspects of prefrontal damage it is executive function, a process that makes use of specific short-term memory stores from the posterior visual and verbal areas to store and search for long-term memories. It is not the memories that are impaired but rather the ability to manipulate and process them: exactly what Baddeley termed the “central executive.”
There is evidence from the human clinical literature that dam-
age to a fairly localized area in the left parietal cortex massively impairs short-term memory of verbal material. A classic example is the case of KF, who was studied by Elizabeth Warrington and Timothy Shallice in England. KF appeared to have relatively normal visual short-term memory but had virtually no verbal digit span. He could repeat back two digits at most. This is strikingly reminiscent of Baddeley’s phonological loop. Other patients with damage to right-hemisphere visual association areas have a dramatic and selective loss of visual short-term memory ability—Baddeley’s visuospatial sketch pad.
But what do the phenomena of short-term visual memory and short-term verbal memory have to do with the prefrontal cortex? Lesions of the human prefrontal cortex do not impair short-term memory processes or, for that matter, many aspects of longer-term memory. Extensive studies by Arthur Shimamura at the University California at Berkeley and by other groups showed that frontal lobe damage seems to impair more the processes than the facts of memory. Thus, such patients are much impaired in the retrieval of information. They have a particular problem with “source” memory, meaning they can remember facts they recently learned, but not where and when they learned them. We saw earlier that this type of memory is termed episodic and refers to events you have experienced, and that this kind of memory is contrasted with semantic memory—for example, your vocabulary—that is not tagged to your own life experiences.
A case can be made that Baddeley’s central executive function is performed by the prefrontal cortex, that the visuospatial sketchpad operates in the posterior visual association cortex, and that the phonological loop may use the short-term verbal memory area in the posterior cortex. In humans (as in monkeys) there are extensive interconnections of these short-term visual and verbal memory areas with regions of prefrontal cortex.
Extensive imaging studies in humans have demonstrated the role of the prefrontal cortex in memory retrieval, a key aspect of working memory. There is an intriguing asymmetry in the retrieval function of the prefrontal cortex. The left prefrontal cortex
seems particularly involved in retrieving semantic information, whereas the right prefrontal cortex is more involved in retrieving episodic information, perhaps consistent with the fact that in most people language functions are located in their left hemispheres.
The frontal lobes have expanded enormously from monkeys and apes to humans. It has been known for many years that people with large lesions of the frontal lobes in both hemispheres have great difficulty inhibiting inappropriate behaviors. The classic case is Phineas Gage, who had a metal rod blown through his skull that obliterated his frontal lobes. He changed from a modest, reliable, hard-working individual to a very erratic and emotional person given to irrational rages (see Box 2-3).
BOX 2-3 One of the most obvious and striking effects of frontal lobe damage in humans is a marked change in social behavior and personality. Perhaps the most publicized example of personality change following frontal lobe lesion is that of Phineas Gage, first reported in 1868. Incidentally, Fred Gage, a recent president of the Society for Neuroscience and a pioneer in the study of new neurons being formed in the adult brain, is a descendant of Phineas. Gage was a dynamite worker and survived an explosion that blasted an iron-tamping bar (3 feet 7 inches long and 1.25 inches wide) through the front of his head. After the accident his behavior changed completely. He had been of average intelligence and was “energetic and persistent in executing all his plans of operation.” After the injury his personality was described as follows: This equilibrium or balance, so to speak, between his intellectual faculties, and animal propensities seems to have been destroyed. He is fitful, indulging at times in the grossest profanity, manifesting but little deference for his fellows, impatient of restraint or advice when it conflicts with his desires, at times pertinaciously obstinate, yet capricious and vacillating, devising many plans of operation, which are no sooner arranged than they are abandoned in turn for others appearing more feasible. A child in his intellectual capacity and manifestations has the animal passions of a strong man. |
But the frontal lobes do not function simply to inhibit bad behavior. We know they play a critical role in overseeing memory; that is, they perform the executive function in memory. When you try to remember something that doesn’t immediately come to mind, the frontal lobes show a marked increase in activity. The memory you are searching for, perhaps something you earlier saw or heard, is not stored in the frontal lobe. Rather, such memories are thought to be stored in cortical areas more toward the back of the brain. But the frontal lobes seem to be the system that is searching through your memory banks. A frustrating aspect of this kind of memory search is called “tip of the tongue.” You know that you know the word or name, but you just can’t remember it at the moment. Sometimes, if you stop trying to remember it and think about something else, the memory suddenly pops into your head, as if during this period the frontal lobes have been busily searching through your vast memory stores until they find the memory you want. As we will see later, such memory searches are not random but rather are guided by associations among the items in memory. This executive function of the frontal lobes in memory is critically important in short-term or working memory. Brain imaging work with normal humans suggests that a major executive function of the frontal lobes in working memory is to inhibit irrelevant or outdated memories. Think of the memory demands on the referee in an ice hockey game. In order to assign credit to players for goals and assists, he must continually keep track of the last two or three players to have control of the puck. Given the nature of hockey, this changes quickly, and the information in the referee’s working memory has to be updated equally quickly.
There is growing evidence that this prefrontal cortex is critically involved in the mental disorder known as schizophrenia. Such patients characteristically have thought disorders, including marked impairment in working memory. Recent evidence, much of it from the work of Patricia Goldman-Rakic, shows that patients with schizophrenia have a significantly reduced metabolic rate in the prefrontal cortex and in fact have a reduced amount of neural tissue in the prefrontal cortex.
Recent studies by Adrian Raine and his associates at the University of Southern California have implicated the prefrontal cortex in violent behavior. These researchers did imaging studies of the brains of normal subjects and convicted murderers. Most of those who had been convicted of murder apparently killed in violent rage, usually one victim and often a relative or friend. But some were mass murderers who killed many times before being caught. Their killings were carefully plotted and not impulsive. Raine and his associates found that impulsive-rage killers had much reduced prefrontal activity compared to normal subjects when performing a task that engages the prefrontal cortex. Interestingly, the mass killers had normal prefrontal activation. It is tempting to conclude that the rage killers had reduced frontal control of impulsive behavior, whereas the mass killers had normal prefrontal function (though clearly they had other abnormalities as well).
Working Memory and Intelligence
Exploring the concept of working memory and its brain substrates has taken us into unexpected territory such as personality, schizophrenia, and criminality. It also takes us directly into one more important area: intelligence. Working memory tasks usually require a person to transform information, to keep track of changes and update memory, to retrieve information, to divide attention, and to make comparisons. If this sounds to you like a description of intelligent behavior, you’re right. Many studies of working memory have shown a close connection between working memory efficiency and general cognitive ability. Correlations of working memory scores and cognitive ability (or IQ) are large and positive, meaning that people with good working memories tend to have above-average cognitive ability as measured by standardized tests (the SAT and GRE, for example). Even performance with the humble memory span test with digit lists is correlated with other cognitive abilities, such as SAT verbal scores. The correlation is not large, but it can be made much larger by a simple
change in the classic digit-span test that requires the subject to repeat the digit list backward. This is a much harder test than the usual digit span test, and the reason seems to be that the backward digit span test requires more working memory resources. You have to mentally manipulate, scan, and reorganize the digit list; you can’t simply hold in it short-term memory and then “dump” it.
Working memory appears to be a special function of the frontal lobes. Is it possible that people who differ in their working memory abilities also differ in the structure or capacity or functioning of their frontal lobes? There are good reasons for thinking this is so. First, there is significant natural variation among people in brain structure, including variation in gray matter volume, which is related to the number of cortical neurons. A large proportion of this variation appears to be heritable and is therefore genetic in origin. In one study of 10 pairs of identical twins, it was found that the correlation of gray matter volume was an astounding +.95, meaning that the two members of each twin pair had essentially identical gray matter volumes. One of the four main dimensions of the Wechsler Adult Intelligence Scale is in fact called Working Memory, and scores on this dimension also have relatively high heritability.
It has not yet been shown that the size of the frontal lobes in different individuals is correlated with working memory performance. Also, at this time, it is not clear that the extent of brain activations is correlated in any simple way with working memory performance. One MRI study found positive correlations between reasoning task scores and frontal lobe activations during problem solving, but some PET studies have found decreased levels of activation in people with high IQs, as if those individuals found the tests easier than subjects with lower IQs. Stay tuned.