Mendel in the Kitchen: A Scientist's View of Genetically Modified Foods (2004)
Chapter: 4 Genes and Species
4
GENES AND SPECIES
The classification of nearly all cultivated plants is now in such a perilous state that the use of any such Latin term as Phaseolus vulgaris is just an elaborate and technical way of saying, “I do not know.”
—Edgar Anderson (1967)
Teosinte is no longer called Euchleana mexicana. Once it was accepted as the ancestor of corn, its genus name, Euchleana, and corn’s genus name, Zea, were lumped into one classification and teosinte became known as a subspecies, called parviglumis, of corn, Zea mays.
Likewise when the goat grass called Aegilops squarrosa was found to have contributed its entire genome to bread wheat, it too lost its name and species classification. Rather than following Edgar Anderson’s whimsical suggestion that the wheat genus be renamed Aegilotriticopyron, to give due credit to its roots, taxonomists decided to demote Aegilops squarrosa and the other 20 or so Aegilops species and put them into the wheat genus, Triticum. Aegilops squarrosa was duly renamed Triticum aegilops. Other taxonomists, however, refused to use the new name (or preferred yet a third name), and international symposia have been convened several times to debate the value of the switch. “The name issue is not yet settled,” argued L. A. Morrison of Oregon State University in 1998. “There is confusion about the number of species in the genus Triticum. There is confusion about the correct names of the species.” Morrison urged other wheat researchers to
“return to the traditional concepts of Triticum and Aegilops as separate genera.”
First teosinte is a species in a different genus, then it is a subspecies of corn. First Aegilops squarrosa is a species in its own distinct genus, separate from wheat, then it is not and its entire genus disappears. Though it might reappear. The plants in that genus are not extinct, of course. Indeed, they have not changed at all—only our way of looking at them has changed. The classifications “genus” and “species” are not fixed and immutable. They are, explains Ernst Mayr, author of Systematics and the Origin of Species, conveniences: they provide “a reliable, easy-to-use, filing system for the bewildering diversity of nature.” C. F. Bessey noted in the American Naturalist in 1908, “Nature produces individuals and nothing more. Species have been invented in order that we may refer to great numbers of individuals collectively.”
The filing of plants into a system of genus and species was begun in 1753 by the Swedish botanist Carl Linnaeus, who was also the first to prove that plants were sexual creatures like animals. The tradition of giving each plant a two-part name (first genus, then species) has held since then, but the rules have changed. Linnaeus, like Aristotle long before him, classified plants and animals chiefly by how they looked. Paleontologists use a similar idea of species to classify fossils. Their “morphospecies” are often based on single fossils. Yet individuals in the same species can differ considerably in size. Should each size of dog be a new species? This definition also fails to account for organisms with very different-looking life stages. Should the caterpillars and the butterflies they turn into belong to different species?
Seeking to include evolution in the definition of species, paleontologist George Gaylord Simpson called a species “a series of ancestor descendent populations passing through time and space independent of other populations, each of which possesses its own evolutionary tendencies and historical fate.” The problem with Simpson’s definition is that there are gaps in fossil records, and such gaps can create arbitrary boundaries between species.
Searching for a better definition of “species,” we simply find more definitions. Ernst Mayr’s biological concept, that “Species are groups of actually or potentially interbreeding populations which are repro-
ductively isolated from other such groups,” is similar to one of the earliest definitions, that of John Ray, who in 1682 defined a species as “a set of individuals who give rise through reproduction to new individuals similar to themselves.” Population biologist Theodosius Dobzhansky early in the twentieth century said that species “are systems of populations; the gene exchange between these systems is limited or prevented in nature by a reproductive isolating mechanism or several such mechanisms.” Yet the mechanisms Dobzhansky had in mind include being on the other side of a mountain, or living in the top of a tree rather than at the bottom. By this definition, people living on different continents in the days before ship travel belonged to different species.
Defining a species as a group of individuals that look alike or can breed together is even more problematic in plants. Indica rice and Japonica rice, for example, are two popular types of the cultivated rice, Oryza sativa. They are members of the same species, and it is often difficult to tell if a single grain comes from one type or the other. While Japonica is usually short-grained and Indica is long-grained, more than a third of the time a grain of Indica is short enough to look like Japonica and a grain of Japonica is long enough to seem to be Indica. Likewise the two types overlap in how they respond to drought, cold, pesticides, and other challenges. Yet they do not crossbreed. When scientists force the crossing—using laboratory techniques very much like those called into play to help an infertile woman bear a child—the resulting Indica-Japonica hybrids are sterile and do not set seed. The two types of rice have been separate for at least 7,000 years. Yet they are still considered the same species.
Then, on the other hand, there are the brassicas. In the grocery store, cabbages, kale, kohlrabi, cauliflower, broccoli, and Brussels sprouts are all neatly divided into their own bins. Diners who ordered Brussels sprouts in a restaurant would be outraged if they were served cabbage instead. And yet all of these common vegetables—even the purple Graffiti cauliflower—are members of the same species, Brassica oleracea.
The original cabbage was apparently native to the Mediterranean coast, where it was grown for its oily seeds, although wild cabbages also

The many faces of Brassica oleracea
grow along the northwestern coasts of Europe. The Greeks were growing leafy kales, probably for livestock feed, by 2,500 years ago. The Romans enjoyed loose-headed cabbages. True hearting cabbages first appeared in Germany, in both white and red varieties, in the 1100s. Cauliflower, kept white by tying its leaves over its head, was being grown in Europe by the 1500s. The advent of kohlrabi, with its swelled stem, is unrecorded, but broccoli, then called “Italian asparagus” or “sprout colli-flower,” began to be popular in the late 1600s. Perhaps people had become more patient by then. As one food historian notes, “I am not certain people realize that many broccolis are at least a year old when they eat them. Like other cabbages, broccolis usually require two years to come to flower, and it is the flower that we are eating.” If allowed to open, he noted, “the flowers would be waxy yellow and fragrant. Butterflies adore them.” Last to the table came Brussels sprouts, showing up as a sport (or mutant) in a Belgian garden in 1750.
If grown carefully, with no chance of sharing pollen, each of these varieties of Brassica oleracea—as well as all their subtypes, such as
purple cauliflower—breeds true: broccoli seeds produce more broccolis, cauliflower seeds more cauliflowers, and so on. But chaos awaits the unwary gardener. Brassicas are outbreeders. They prefer—they are “anxious,” say the botanists—to mate with plants that are genetically unlike themselves. Some are even self-incompatible, meaning that they will not use their own pollen to fertilize their own flowers even if no other pollen is available. (Apple trees are self-incompatible too, which is why orchardists plant different varieties in alternating rows.) In an experiment in England, botanist W. F. Giles planted a mix of garden-variety cabbages and kales in a square and allowed them to cross-pollinate. “The work of centuries was wiped out in a generation,” reports one observer. The cabbage heads disappeared and the wide, branching, gangly look of the wild plant took over.
And the promiscuous Brassica oleracea tribe will not only mate with each other. Some variety of cabbage in seventeenth-century Bohemia crossed with a turnip, which is in the genus Brassica, but a different species. This cabbage-turnip cross is the rutabaga. Rutabaga is classified as a variety of Brassica napus. Another variety of that species is now widely grown—like the original domesticated cabbage—for its oily seeds, the source of the popular canola oil.
![]()
The danger of crossing the species barrier—even the immorality of it—is often cited as a reason to restrict, or even prohibit, the molecular modification of plants. In 1984 activist and writer Jeremy Rifkin testified before Congress that transferring a gene from one species to another represents “a fundamental assault on the principle of species integrity.” In 1998 the authors of Genetically Engineered Foods: Are They Safe? You Decide wrote that “The mixing of genes by crossbreeding is clearly subject to very definite rules—you can’t mix unrelated species…. Where there are rules there are boundaries…. Natural law has set a boundary. Genetic engineering is not constrained by these rules and crosses all boundaries set in place by natural law.” To support their case, they quote John Hagelin, a physicist: “When genetic engineers disregard the reproductive boundaries set in place by natural law, they run the risk of destroying our genetic encyclopedia, compromising the richness of our natural biodiversity and creat-
ing ‘genetic soup.’ What this means to the future of our ecosystem, no one knows.”
In 2000 Representative Kucinich argued in Congress that “Conventional breeders are bound by species boundaries that allow them to transfer genetic material only between related or closely related species. By contrast, the very purpose of genetic engineering is to allow scientists to transfer genes from completely unrelated life forms.” In 2001 journalist Bill Lambrecht defined “genetically modified organism” in Dinner at the New Gene Café as “what you get when you move genes across the traditional species boundaries of plants and animals in the quest for new traits.”
These critics are apparently operating under yet another understanding of the word “species.” One such definition comes from the Middle Ages, when scholars rediscovered the works of Plato and Aristotle and reinterpreted them in a way that had a lasting effect. Johnny Appleseed’s Swedenborgian notion that every apple tree was a reflection of a spiritual tree, and therefore should not be grafted or otherwise kept from expressing itself, has its source in this medieval worldview. According to Thomas Aquinas, the thirteenth-century Italian philosopher, everything on Earth is but a reflection (albeit a poor reflection) of what exists in Heaven. Each species represents an idea in the mind of God.
This medieval concept of species was a major stumbling block for the theory of evolution when it was first introduced in the mid-nineteenth century. The French scientist Jean-Baptiste Lamarck set forth in 1809 the idea that creatures had changed—or evolved—since God created Earth. This idea seemed to explain satisfactorily how a cabbage could become a broccoli. But Darwin, in The Origin of Species, went further. He argued that one primordial plant could branch into many lineages, giving rise to all flowering plants, from broccoli to apple to daffodil. (Not to mention that one primordial ape could give rise to both monkeys and humans.)
Even scientists who did not think Darwin’s argument was blasphemous thought they saw a flaw in his reasoning. Darwin’s theory of natural selection assumes that individuals within a species differ in how fit they are. Those whose fitness best matches the challenges of
their home environment survive to have more children and pass down to those children something that will also, by and large, make them more fit. The accumulation of small improvements in fitness, generation by generation, eventually leads to the branching off of a new and better species.
The problem lay in that first assumption. Everyone agreed that individuals were different. But were those differences inherited? On what did natural selection act? “Not until the 1930s was this missing element filled in,” notes Harvard physiologist Bernard D. Davis. That element was the gene. To know what species—or species barrier—means to a modern-day plant scientist, to see what Golden Rice, triticale, bread wheat, broccoli, hybrid corn, and canola oil might have in common, takes an understanding of the gene.
Like the notion of a species, the concept of the gene went through many revisions before a scientific consensus was reached and, like species, it has changed dramatically since then, as we have learned more about what actually occurs inside a cell.
The field of genetics, the science of heredity, began in 1900 when Hugo De Vries and Carl Correns rediscovered the pea experiments Mendel had done in his monastery garden half a century earlier. Correns wrote, “The same thing happened to me which now seems to be happening to De Vries: I thought that I had found something new. But then I convinced myself that the Abbot Gregor Mendel in Brünn, had, during the sixties, not only obtained the same result through extensive experiments with peas, which lasted for many years, as did De Vries and I, but had also given exactly the same explanation, as far as that was possible in 1866.”
In Darwin’s and Mendel’s time, the mechanics of inheritance were not well understood. Most people in the nineteenth century thought of it as a blending of traits, like a mixing of water and wine or two colors of paint. What Mendel found was far from blending and so did not seem to relate to ordinary inheritance. Mendel crossed pea plants that bore round seeds with pea plants that bore wrinkled seeds. When he planted the seeds of the cross-pollinated plants, the next generation (called the first filial, or F1, generation) bore only round seeds. The seeds were not only less wrinkled than their wrinkly parent (as they
might have been if the two traits had blended), they had no wrinkles at all. The trait had disappeared. But curiously, when these F1 plants were allowed to pollinate themselves (which peas prefer to do, being natural selfers, not anxious outbreeders like the brassicas), they produced round and wrinkled seeds in a ratio of three round to one wrinkled. The trait that had vanished in the F1 generation reemerged unchanged in the F2 generation.
Mendel showed that this behavior was simple, regular, and reproducible. His explanation was simple, elegant—and revolutionary. Traits, or as he called them, “differentiating characters,” underwent no blending at all. Each organism had two hereditary units for each trait, one from the father and one from the mother. Only one of these two units would determine the trait. This one was dominant. The other, the one that vanished in the F1 generation, he called recessive. If a seed that received both a dominant (roundness) unit and a recessive (wrinkliness) unit was round in appearance, then the three-to-one ratio was easily explained. Because half of the parents’ hereditary units called for round seeds and half called for wrinkled seeds, one quarter (half of a half) of the offspring would receive only roundness units and another quarter would receive only wrinkliness units. The rest would receive a roundness unit from one parent and a wrinkliness unit from the other parent. Because roundness obscures wrinkliness, three-quarters of the seeds would be round and only one-quarter—that quarter receiving two wrinkliness units—would be wrinkled.
These hereditary units did not change even when they could not be seen. Nor did the inheritance of one trait seem to affect the inheritance of another. When Mendel bred not only round and wrinkled peas, but green and yellow ones, he found that the two traits “assorted” independently. Only 1 in 16 (a quarter of a quarter) of the F2 offspring were both wrinkly and yellow, meaning that they had two recessive units for each trait. But while Mendel could predict how many of the plants in his garden each season would bear yellow peas, he could not explain how the process worked.
As the field of genetics began to develop in the twentieth century, first the units of heredity were named, then they began to acquire a physical identity. De Vries, who had coined “mutation,” decided to call
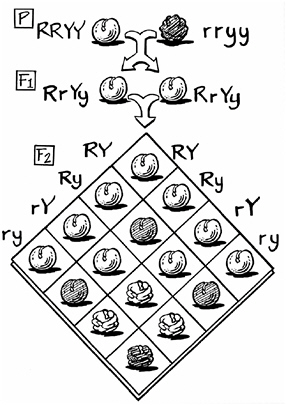
Mendel’s experiment with peas, showing the assortment of round (R), wrinkly (r), yellow (Y), and green (y) traits
them pangenes, from Darwin’s concept of pangenesis, which said that every cell in the organism sent a tiny bit of itself to the egg or sperm so that the resulting embryo would know how to recreate a whole organism. Other geneticists came up with the names plasomes, idioblasts, and biophores for the same particles. Each name had an earlier theory attached to it, and none of them satisfied everyone. Then in 1909 Danish botanist Wilhelm Johannsen published his work on beans. “It appears simplest to isolate the last syllable, ‘gene,’” he wrote, from De Vries’s pangene. “The word ‘gene,’” he noted, “is completely free from any hypotheses.” The term stuck, as did Johannsen’s definitions of phenotype, for the outward appearance or character of an organism (in Mendel’s peas, wrinkled or round), and genotype, for the combination of gene variants (or alleles) the organism carried: either two dominant types, two recessive types, or one of each.
Still, it took a while before scientists agreed that these particles, these genes, were things—that they had physical form that could be
observed and changed. In 1912 Edmund M. East, the expert on corn genetics who thought hybrid corn was a waste of effort and believed teosinte could not possibly be the ancestor of corn, thought the gene was only a useful shorthand. “As I understand Mendelism, it is a concept pure and simple,” he wrote, “a conceptual notation as is used in algebra or chemistry. No one objects to expressing a circle as x2 + y2 = r2.”
Two years later Thomas Hunt Morgan at Columbia University would claim that a gene was “some minute particle of a chromosome whose presence in the cell influences the physiological processes that go on in the cell.” Chromosomes had long been observed in the cells of both plants and animals. In 1875 the German botanist E. Strasburger had described spindly structures that, as the cell divides, first lengthen, then shorten again to form compact little rods. In 1888 botanist Wilhelm Waldeyer noticed that these rods took up a stain well, so he named them “chromo” (colored) “somes” (bodies). But no one had, as yet, connected them to genes.
Morgan, who would do so, began graduate school at Johns Hopkins University in 1886. There he was taught by a self-proclaimed Darwinist, William Keith Brooks, who had worked with William Bateson, the geneticist who later introduced Mendel’s laws to English science. Brooks tried to interest Morgan, as he did all of his students, in the study of heredity. But after completing a dissertation that used a comparison of embryos to prove that sea spiders were true spiders, not crustaceans like lobsters, Morgan went to Italy to continue studying how eggs develop.
One theory at the time suggested that within each egg was a tiny manikin, a miniature creature or homunculus, that could somehow unfold into an adult. To test this theory, Morgan took a two-celled embryo of a whitefish and destroyed one cell. The remaining cell grew into an “incomplete” adult, he found—unless he turned it upside down, shook it, or whirled it in a centrifuge. Then it developed normally. Clearly things were more complicated than the manikin theory supposed.
In 1903 Morgan went to Columbia University as America’s first professor of experimental zoology. Inspired by Mendel’s laws of hered-
ity and De Vries’s mutation theory, he experimented on all sorts of animals. He used cockroaches, crabs, tadpoles, sea urchins, starfish, earthworms, flatworms, jellyfish, mice—by one estimate, more than 50 different creatures—before he settled on the fruit fly, Drosophila. And unlike his predecessors, who had contented themselves with observing the bewildering variety of nature, Morgan and his students induced mutations, creating odd-looking flies (yellow bodies, white eyes, pink eyes, tiny wings) by exposing them to heat, cold, salt, sugars, acids, alkalis, radium, and X-rays.
His laboratory was known as the Fly Room. It “was a rather small room,” recalled Alfred Sturtevant, “with eight desks crowded into it, in which the three of us reared Drosophila for the next 17 years.” Sturtevant joined the Fly Room in 1909. An undergraduate, a teenager, he had impressed Morgan with a paper on the inheritance of coat color in his family’s stable of racehorses. He, in his turn, was impressed by a lecture in which Morgan described what he called the strength of coupling or linkage of certain genes. Strangely, these fruit fly genes did not assort independently, as had the genes of Mendel’s peas. Some appeared to stick together—to be coupled to each other.
In Mendel’s peas, the fact that an offspring received one trait (for instance, wrinkly seeds) made it no more or less likely that it would receive a second trait (green seeds). But in Morgan’s fruit flies, some traits tended to be inherited together. If a fruit fly offspring had white eyes, it was also likely to have a yellow body. The reason, Morgan believed, had to do with where the genes physically resided in the cell. Genes, he argued in his book The Mechanism of Mendelian Heredity, were not abstract concepts. They were physical units, located at definite positions on the chromosomes. Genes that were linked must be near each other on the same chromosome. The closer they were, the more likely the two traits would be inherited together. The farther apart they were, the more likely it was that they would not be inherited together. The reason has to do with the way chromosomes exchange parts or recombine during meiosis, when the sex cells are formed.
It was Morgan who named such recombination “crossing over.” Crossing over happens just before the chromosomes split into two haploid, or half sets, each of which furnishes one normal egg or sperm
with its genetic material. Each DNA helix in the chromosome is cut in several places and a bit is chewed off each end. The loose ends invade their counterpart chromosome, the homolog (from the Greek for “agreeing in proportion”) received from the other parent, pushing out one strand of its DNA. The homologs exchange parts, each carrying genes from the previous generation. Then the DNA strands in the chromosomes are reconnected. Some genes received from the grandmother are now attached to genes received from the grandfather. In this way, genes from grandmother and grandfather are mixed before being passed down to the grandchild. Genes that are linked are located on the same cut bit of chromosome, and so are passed down together. Sturtevant was intrigued by Morgan’s idea. He talked his way into Morgan’s lab and devised a test for his hypothesis. Like Morgan,
Sturtevant and another student, C. B. Bridges, kept their mutant flies in hundreds of half-pint milk bottles, feeding them mashed bananas. The milk bottles were hardly a high biosafety-level containment facility for these genetic experiments. “Near the entrance of the room a stalk of bananas hung conspicuously, serving as a center of attraction for the numerous fruit flies that had escaped from their milk bottles or that had bred themselves, without the benefit and direction of science, in the garbage can that was never thoroughly cleaned,” wrote Ian Shine and Sylvia Wrobel in their biography of Morgan. The lab window was often left open, both to lure in flies that could then become breeding subjects and because the room stank of fermented banana. Morgan had other experiments going on in the lab, including some “whose purpose no one ever figured out exactly, like the one in which a crab walked around with another crab glued to its back, a fragment of radium between the pair.”
By tracking fruit fly mutations over generations, Sturtevant was able to draw the first genetic map. Published in 1913, it showed six genes—those for yellow bodies, white eyes, vermilion eyes, miniature wings, and rudimentary wings—in a straight line on the fruit fly’s sex chromosome. Morgan’s definition of a gene seemed correct. A gene was a specific, fixed point on a chromosome: a locus, from the Latin for “place.” Genes were like beads on a string, like pearls in a necklace, like dots on a line. The gene had been defined as the unit of heredity.
![]()
It soon became obvious that the number of linkage groups in a fruit fly, a pea, or a person was the same as the number of chromosomes in a cell. Knowing this, scientists were able to tie each linked group of genes to a chromosome. By 1936 corn breeders, for instance, could make use of a map that located 100 corn genes onto specific chromosomes. They could track the pattern of inheritance of another 250 genes from crop to crop.
Each bead on the string, each gene, directed the production of one enzyme, George Beadle and Edward Tatum at Caltech determined in 1941 after studying mutations in orange breadmold. (Later Linus Pauling, at the same institution, would expand their idea: each gene controlled production of one protein, he said, of which enzymes are only one kind). In 1944 Oswald Avery, Colin MacLeod, and Maclyn McCarty at the Rockefeller Institute for Medical Research in New York carried out experiments that led to the identification of the chemical nature of genes. They found that a chemical called deoxyribonucleic acid, or DNA, extracted from a virulent bacterium, could by itself transform a harmless form of pneumonia bacteria into a virulent type when the two were injected into the same mouse.
Until then DNA had been dismissed as a “stupid” molecule, in the words of physicist Max Delbrück. It was thought to be too simple to be of much biological importance. It was made up of just four small molecules, the nucleotides, each of which had a sugar, a nitrogenous base, and a phosphate group. Nor were the bases themselves of much interest. Adenine (now known familiarly as A) was common in beef pancreas. Cytosine and thymine (C and T) were purified from the thymus glands of calves. Guanine (G) had first been found in guano (bird feces); in crystallized form, it gave the shine to fish scales.
Proteins, on the other hand, had long been thought special. Discovered in the 1830s, they had been named from the Greek proteios, meaning “of the first importance.” They were long chains or polymers made up of 20 different small molecules, the amino acids. Proteins might be complex enough to make a gene, but certainly DNA was not. Then in 1952 Al Hershey and Martha Chase showed that when a virus called a bacteriophage infects a bacterial cell, it injects only its DNA
into the cell, abandoning its protein coat. Once again, the active ingredient was DNA. Suddenly DNA was the “molecule of life.”
How could complex genetic information—the making of a white eye or a wrinkled pea—be conveyed by a simple molecule with only four constituents, A, T, G, and C? The secret lies in how those parts are assembled—both in the order of the letters and in the structure of the DNA molecule. As James Watson and Francis Crick announced in a short paper in Nature in 1953, DNA is a double helix, rather like a spiral staircase. Watson and Crick had earlier proposed to their London colleagues, Rosalind Franklin and Maurice Wilkins, that DNA was a triple helix. Franklin, however, pointed out that they had mistaken (by a factor of 10) the proportion of water in the structure. Franklin was a crystallographer. She took photos of the diffraction patterns created when crystals of DNA were exposed to X-rays. Studying the patterns, she was learning about the molecule’s structure. She had not yet published her photos, or fully interpreted what they meant, when Wilkins arranged for Watson to look at the best of them. In 1962 Watson, Crick, and Wilkins received the Nobel Prize in physiology or medicine for discovering the double helix. (Franklin died of cancer before the prize was awarded.)
“The novel feature of the structure,” Watson and Crick wrote of the double helix, “is the manner in which the two chains are held together.” Because A can pair only with T, and G can pair only with C, the two chains are mirror images of each other. “It has not escaped our notice,” Watson and Crick wrote in the paper’s final statement—or understatement—“that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.”
By 1969 the DNA code was declared cracked. Each sequence of three letters, each codon, such as GAG or GAA or AAA, stands for an amino acid (except for three codons, which instead mean “stop”). When the code is read, the cell’s machinery links the specified amino acids together into a chain, which is then folded up to become a functioning protein. Each protein made by the body has its corresponding DNA code—a gene. (Scientists generally say the gene encodes, or codes for, the protein.) Each bead on the string, each gene, turned out to be
more like a sentence written in three-letter words. Each sentence was the recipe for a protein. Mendel’s “differentiating characters” could thus be explained as alternative forms of the same gene: one codes for a normal protein; the other does not because one or more words in the sentence had changed, or mutated.
The genome—the sum total of an organism’s DNA—was understood to be its book of life, life’s little instruction book. The analogy is, of course, not exact. A true book contains many pages of sentences folded in upon each other: you read across a line of letters, then go to the beginning of the next line. To turn a true book into the book of life, you must take out the spaces between words. Connect the last letter on each line with the first letter of the first word on the next line until you have strings of letters folded one upon the next. Then grab the last word in the book and pull—the letters of the book will stretch on for miles. So it is with DNA.
This long string of meaning, made up of words and punctuation all fashioned from the same four letters, A, T, G, and C, is folded up into the chromosomes of a cell. How to read it—how to learn the sequence in which the four letters are linked—was discovered by two teams of scientists in the late 1970s. First they divided the chromosomes into parts—like unbinding a book so that its pages are loose. Then they made several copies of one part—one page—at a time. These copies they cut randomly into different lengths. Knowing the length of each fragment and what its last letter was, they could decipher the order of the letters. If the code were written in English, for example, they might have had a four-letter fragment that ends with “e,” a two-letter and a five-letter fragment that both end with “r,” and a three-letter fragment that ends with “d.” Given a one-letter fragment that is an “o,” the word is “order.”
Today genomes are being sequenced more rapidly than they can be analyzed. Much of the process is automated. Automated, too, is the comparison between one genome and another. Still, each genome hides its secrets. Although we know its code—how the letters of its ATGC alphabet are translated into proteins—we cannot read it like a book. The analogy to a book is, in the end, inadequate, because what is written in the DNA is how to make proteins (and other molecules, like RNA), not how to make an organism.
When the first complete draft of the human genome sequence was published in the journals Science and Nature in February 2001, Science lead author Craig Venter wrote of “a major surprise.” There were far fewer genes—no more than 30,000 or 40,000—than the 50,000 to more than 140,000 that had been predicted. That’s hardly more genes than the 26,000 in the genome of Arabidopsis thaliana, the little weed used as the fruit fly of plant genetics. Drosophila itself has 13,000 genes. Venter concluded: “The modest number of human genes means that we must look elsewhere for the mechanisms that generate the complexities inherent in human development.”
Nor are there only genes in the genome, if by gene we mean the sequence that codes for a protein. Of the string of 3.2 billion ATGCs that make up the human genome, for example, only a tiny fraction—between 1.1 and 1.4 percent—actually codes for the organism’s proteins. The coding sequences are scattered throughout a much longer length of DNA in short segments called exons. They are interrupted by noncoding segments, introns, that can be tens of thousands of letters long. The functions of introns are still being discovered, though they are now generally considered to be part of the gene. Introns can increase the transcription of a gene; that is, genes work better with introns.
There are other sequences called promoters and enhancers. These regulate the genes, specifying when a gene is turned on and how active it is. Some of the DNA codes for molecules that guide the editing of gene transcripts—the splicing together of exons and the excision of introns. Other sequences code for regulators or have a structural function. Yet even counting the introns as part of a gene, less than 30 percent of the genome is devoted to genes.
The rest was once labeled “junk” DNA. The name stuck, even after the nature of these sequences was understood. One type of sequence was originally called satellite DNA because it had a rather different composition than the mainstream DNA and therefore formed a satellite band in a standard laboratory analytical procedure. Today such sequences are known to consist of many, many copies of very short sequences and to be important for cell division: they apportion chromosomes when cells divide. But much of what was originally labeled “junk” is made up of transposons, popularly known as jumping genes.
Transposons are genes that can move from one place to another on the chromosome. Although the patterns they form in plant leaves and flowers were well known, it was Barbara McClintock’s studies in the colorful maize called “Indian corn” that led to the first reports, back in the late 1940s, that such patterns were caused by genes that move. They remained an abstract concept, like the early twentieth-century notion of the gene itself, until the first one was isolated and cloned decades later. Since then, they have come to explain quite a lot about how genes act—and evolve.
For instance, in 1990 scientists identified the gene that had mutated to give Mendel’s peas their wrinkled seed trait. The gene codes for a protein that helps make starch in the seed. The mutation was caused by a transposon: by jumping into the gene’s sequence, the transposon made it impossible for the complete protein to be made. Without this protein, less of the sugar in the pea can be converted into starch. With less starch, the pea seed wrinkles as it dries. And in the early 1980s, it was an understanding of transposons—what they are and what they can do—that inspired scientists to harness the soil bacterium Agrobacterium tumefaciens to carry new genes into plants.